Any extracts used in the following article are for non-commercial research and educational purposes only and may be subject to copyright from their respective owners.
ChatGPT or AI bots played no part in the generation of content, and never will.
Reading time estimate: 2h 19m
Last updated:
12th February ‘23: Further NEAT1 & MALAT1 related pathologies.
25th February ‘23: Epigenetics & prionopathies.
10th March ‘23: Link added to the Substack on therapeutic interactions.
Credit must go to Dr Annelise Bocquet for bringing attention to a fundamental area of RNA biology which relates to pathologies and immunological responses to viral infections and many other conditions including vaccination and to experimental mRNA gene therapies.
Its an area that has received almost no attention during the last three years of “pandemic” response and “vaccine” development, being dwarfed instead by talk of the much more familiar aspects of immunology such as antibodies, antibodies, antibodies, and maybe even mention of T cells if you were lucky.
This Substack aims to help bridge the knowledge gap to some degree. It is, however and somewhat ironically a huge subject area so I will need to keep within scope as far as possible.
One of the biggest challenges when writing about lncRNAs is what to leave out. Its as if you have a galaxy available to study and you must focus on just a few stars. Meanwhile the vaccine developers aren't even looking at the sky.
What are long non-coding RNA’s (lncRNA)?
LncRNAs are a type of RNA generally defined as transcripts of more than 200 nucleotides that are classically considered to not be translated into proteins1, although there is evidence that some peptides may be expressed, but these are considered highly unstable and without biological function2.
LncRNAs are in a distinct class to small non-coding RNAs that include microRNAs, small interfering RNAs, Piwi-interacting RNAs, short RNAs and small nuclear RNAs.
Only about 20% of the human genome consists of protein-coding genes, the rest being non-coding sequences3. These number at least 270,0004.
Tissue specificity is a notable feature:
Quantitatively, lncRNAs demonstrate ~10-fold lower abundance than mRNAs, which is explained by higher cell-to-cell variation of expression levels of lncRNA genes in the individual cells, when compared to protein-coding genes. In general, the majority (~78%) of lncRNAs are characterized as tissue-specific, as opposed to only ~19% of mRNAs. In addition to higher tissue specificity, lncRNAs are characterized by higher developmental stage specificity, and cell subtype specificity in tissues such as human neocortex and other parts of the brain, regulating correct brain development and function. In 2018, a comprehensive integration of lncRNAs from existing databases, published literature and novel RNA assemblies based on RNA-seq data analysis, revealed that there are 270,044 lncRNA transcripts in humans.
Figure 1.Mechanisms of action of lncRNA-mediated mRNA stability regulation. LncRNAs can modulate mRNA stability through different mechanisms: (A) Direct interaction with miRNA or RBP binding sites in target mRNA; (B) Sequestration of miRNAs or RBPs to avoid their interaction with mRNA molecules; (C) Acting as scaffolds to enhance RBP-mRNA interactions; (D) Interaction with m6A machinery to modulate m6A levels of target mRNAs. https://www.mdpi.com/2311-553X/7/1/3
In regulation of DNA replication timing and chromosome stability.
In aging and disease:
Neurological disease9 and cancer. For example Alzheimer’s disease, prostate tumours, breast cancer, non-small cell lung cancer, oral squamous cell carcinoma and colorectal cancer10.
Significant lncRNAs associated with cancer include MALAT1 (=NEAT2)11, which will be discussed later.
Significant lncRNAs associated with and predictive of heart failure and cardiovascular disease (CVD)include MIAT12, ANRIL13 and HEAT2 or KCNQ1OT11415 .
As the structure of the transcriptome (ie the set of all RNA transcripts, including coding and non-coding, in an individual or a population of cells16) is being interpreted more and more previously unknown associations with disease are being discovered.
Single nucleotide polymorphisms (SNPs)17 associated with disease states have been mapped back to lncRNAs, such as for susceptibility to myocardial infarction (MI)18, coronary artery disease (CAD)19 and atherosclerosis2021.
In vaccines:
LncRNAs are involved in the immune response to vaccination, including for influenza and for yellow fever.
This paper by de Lima et al (2019) discusses their interactions22:
Long noncoding RNAs (lncRNAs) are known to be involved in several immunological processes. In spite of their general relevance to human immunity, to date there are no reports on the importance of lncRNAs in vaccine responses. Here we apply a “systems vaccinology” framework to study the role of lncRNAs in vaccine-mediated immunity. We applied meta-analytical approaches using public microarray data from over 2,000 blood transcriptome samples of vaccinees and an RNA-sequencing (RNA-seq) dataset from vaccinated children to tackle this question. Our results indicate that lncRNAs are important players in several immunological processes elicited by vaccination.
They used microarray platforms to find blood and peripheral blood mononuclear cell (PBMC) expression data from 17 cohorts of human participants immunized with either yellow fever vaccine (YF-17D) or inactivated influenza vaccine (IV)
lncRNAs Present Coherent Changes of Expression after Vaccination.
Dozens of lncRNAs were identified whose expression was induced or repressed after 1, 3, 7, or 14 d following IV compared with baseline (Fig. 2A and SI Appendix, Fig. S3). Among the protein-coding genes, TNFRSF17, GGH, and CD38 were up-regulated 7 d postvaccination in most cohorts (Fig. 2 B and C), as previously reported (6)
Fig.2. Transcriptome analysis of cohorts immunized with inactivated IV. Cumulative sum of differentially expressed lncRNAs (A) and protein-coding genes (B) (y-axis) in one or more datasets (x-axis). Transcripts were considered differentially expressed if limma P values were lower than 0.05 in at least three cohorts (for lncRNAs) or four cohorts (for protein-coding genes). (C) Forest plots of representative lncRNAs and TNFRSF17 with reiterated differential expression. Log2 fold changes with their corresponding 95% confidence interval (x-axis) are plotted for each cohort (y-axis). Red vertical lines and shaded regions represent log2 fold-change summaries and their 95% confidence intervals, respectively. (D) Heatmap depicting gene expression of human immune cells from ref. 28. Logs (FPKM) of genes are scaled around zero. Columns represent samples; rows represent genes. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6708379/
They then performed further analysis on different leukocyte classes and included a cohort of children with the yellow fever vaccine assessment.
T cell functionality may well be affected by lncRNA PRKCQ-AS1 down-regulation, and its subnetwork of genes:
Meticulous examination of genes connecting to these lncRNAs may unveil potential transregulatory relationships (see SI Appendix). For instance, lncRNA PRKCQ-AS1 is a member of the T cell–related community CM5 in the IV network (Fig. 4A). GSEA revealed that CM5 is consistently down-regulated on day 1 after IV in all IV cohorts (Fig. 4C). A subnetwork of genes connected to PRKCQ-AS1 is also broadly down-regulated on day 1 (Fig. 4D). Several members of this subnetwork are well-known T cell–related genes, such as CCR7, CD27, ITK, LEF1, THEMIS, and SKAP1. These findings suggest that PRKCQ-AS1 may be involved with T cell functions.
From analysis of the new paediatric vaccine cohort:
As RNA-seq analyzes the whole transcriptome, it has the ability to discover lncRNAs associated with vaccination. We assessed whether lncRNAs were differentially expressed in an RNA-seq dataset of 26 children aged 2–4 y old immunized with intranasal LAIV. Vaccination induced differential expression of 55 lncRNAs in whole blood at day 2 post-LAIV compared with baseline, with a false discovery rate (FDR) threshold of 0.05 (SI Appendix, Fig. S7). Several lncRNAs found in our meta-analyses were also found among differentially expressed genes in this cohort, including FAM225A, LRRC75A-AS1, MAPKAPK5-AS1, DANCR, and DICER-AS1.
They found that the yellow fever vaccine tended to skew towards antiviral interferon gamma responses (IFN), whereas the influenza vaccine skewed towards antibody-secreting B cells.
Type 1 interferon (IFN-1) is the first defence line of the host antiviral response in the unvaccinated, so although B cell antibodies may inhibit influenza viruses this is usually at a later stage in the infection and would be suboptimal as the first line response23:
Our analyses have revealed that lncRNAs may be linked to some of these processes. While YF-17D contains a live attenuated yellow fever virus that needs to replicate before priming the immune system of naïve individuals, IV contains proteins of the influenza virus that boost a preexisting immunity. At day 7 postvaccination, the blood signature of YF-17D vaccinees is related to innate antiviral IFN responses (3), whereas the signature of IV vaccines is more associated with antibody-secreting B cells (6, 7). These differences may explain the distinct set of lncRNAs associated with each vaccine.
One of the lncRNAs called MIAT is associated with myocardial infarction and oncogenesis. Skewing expression either up or down could promote various pathologies. Downregulation of MIAT is associated with decreased risk of oncogenesis24, but DANCR and LINC01133 were increased. This implies an increased cancer risk for a time, including in young vaccinees.
Hopefully the duration is short enough not to be of concern, but if you are in remission the risk of initiating a recurrence must be considered, and the grade of cancer may be increased for existing patients:
LncRNA transcripts whose functions have been described in other contexts, such as MIAT, LINC01133, and DANCR, were found to be consistently differentially expressed in multiple vaccination cohorts. MIAT, also known as Gomafu, was first described in 2006 as a transcript harboring SNPs conferring increased susceptibility to myocardial infarction (35). More recent reports have shown that MIAT might be involved in processes related to apoptosis, proliferation, oncogenesis, and cellular migration by several mechanisms (reviewed in ref. 36). Consistent down-regulation 1 d after IV across multiple cohorts may point to an unappreciated role for MIAT in the early immune response to vaccines. LINC01133 expression was found to be increased in several cancers (37–39). In our analysis, LINC01133 was found to be up-regulated at day 3 after IV, which suggests that it may also act to regulate immune cell proliferation following vaccination.A role for lncRNAs in adaptive immune responses also needs consideration.For example, DANCR, which was also up-regulated in young vaccine responders at day 7, as discussed in a previous publication (12), was found to influence the activity of Enhancer of Zeste Homolog 2 (EZH2) in multiple cancer types (29, 30, 40). Epigenetic modifications mediated by EZH2 have been shown to affect B cell differentiation and antibody production in mice (41). DANCR up-regulation 7 d after IV suggests that it takes part in regulating the differentiation of antibody-secreting cells, whose frequency in peripheral blood rises at roughly 7 d after IV (11, 13).
Long range DNA interactions between immunoglobulin (Ig) segments appear related to lncRNA expression changes, such as to FAM30A. Class switching may well be related to this too. From a murine study:
Antisense noncoding expression within murine loci poised for recombination has been reported in mice (45–47). Although conclusive functional characterizations of these mouse lncRNAs are still lacking, they were proposed to act in the regulation of variable (diversity) joining [V(D)J] recombination by mediating long-range DNA interactions between Ig segments (46). Network analysis also revealed that FAM30A is a member of CM7, a community associated with B cell–related genes (Fig. 4B and SI Appendix, Table S2). Altogether, our results indicate that FAM30A is probably related to the biology of B lymphocytes and to antibody responses elicited by IV. Although FAM30A is not conserved in mice, therefore preventing murine studies, its association with immune responses to vaccination warrants further investigation.
Vaccine development to date appears to have proceeded without any analysis (or publication) of changes to lncRNAs and the possible unintended pathologies these can introduce, including suboptimal immune responses.
The great thing here is that the authors provided an online database so that other researchers can check for themselves:
This was 2019 and I suspect the researchers were as shocked as anyone reading this at the state of ignorance, wilful or otherwise, and felt compelled to do something:
To the best of our knowledge, there is no study focusing on the role of lncRNAs during vaccine-induced immunity. The work presented herein implicates lncRNAs in the regulation of specific processes associated with vaccination. We have also created an online database where users can submit personalized queries for coding genes and lncRNAs to visualize the results from our meta-analyses.We hope that this comprehensive resource will aid researchers in rapidly assessing hypotheses related to the blood transcriptomics of human vaccines.
MALAT1 suppresses immunity to infection
From 2020, Hewitson et al performed a study using mice, in vitro activation of CD4+ T cells, quantitative RT-PCR, RNA-seq analysis and western blotting25.
MALAT1 upregulation suppresses CD4+ T cell activation and increases IL-10 expression in Th cells, an anti-inflammatory cytokine26.
Again, more research is urgently required, especially if more point-of-care mRNA therapeutics are to be delivered without any long term clinical data:
New medicines with very short shelf lives and highly personalised medicines can more easily be made in or near a hospital setting or ambulance, meaning patients who need them can access them faster.
Despite remarkable progress in mapping lncRNAs to mammalian genomes and exploring lncRNA function at the molecular level in cellular systems, there is a profound lack of understanding of the function of lncRNAs (requirement, sufficiency, or redundancy) at the whole-organism level. For example, although CD4+ Th cells are central to pathogen-specific adaptive immunity (3), and there are hundreds of lncRNAs identified as differentially regulated during CD4+ T cell activation in humans and mice (4–6), fewer than a handful of lncRNAs have been shown to affect Th cell function. These include NeST (7), which has been shown to control its neighboring Ifng locus, and lincR-Ccr2-5′ AS (5) and linc-Maf-4 (6), which affect CD4+ T cell gene expression through long-range interactions. Therefore, the functional relevance of lncRNAs in vivo is a largely unexplored and emerging challenge in both the fields of immunology and RNA biology.
Metastasis-associated lung adenocarcinoma transcript 1 (Malat1) is a 7.5-kb-long long intergenic noncoding RNA (lincRNA) transcript, which is associated with cancer progression and metastasis (8). It is localized in nuclear speckles (9), which are nuclear foci enriched in factors involved in pre-mRNA splicing and transcription (10).
IL-17 is associated with various autoimmune disorders, inflammation and cancer metastasis. MALAT1 expression correlates positively with TH-17 and IL-17:
Having found that Malat1 suppression is a hallmark of Th activation, we tested the effect of Malat1 deletion on Th activation. We found that following in vitro differentiation of naive CD4+ T cells to Th1, Malat1−/− cells displayed a reduction in levels of IFN-γ that did not reach statistical significance but significantly reduced expression of the immunoregulatory cytokine IL-10. Upon Th2 differentiation, there was also a significant reduction in IL-10 levels, with IL-4 being unaffected (Fig. 3A–C). The effect on IL-10 was more prominent in Th2 cells, which express higher levels of IL-10 than Th1 cells in vitro (Fig. 3A, 3B). We also observed a reduction in IL-10 mRNA levels (Fig. 3D). We repeated these experiments under weakly polarizing conditions and found a statistically significant reduction upon Malat1 loss on IFN-γ expression in suboptimally activated Th1 cells but no effects on IL-10 or IL-4 and Il-10 under weakly polarizing Th2 conditions (Supplemental Fig. 3A). Malat1 loss also suppressed IL-10 and IL-17 expression under Th17-differentiation conditions.
Malat1 expression inversely correlates with expression of transcriptional units involved in RNA processing and translation, protein degradation, metabolism, and cellular structure, all hallmarks of Th activation. Similar correlations are seen in Th2 cells. Conversely, Malat1 expression positively correlates with expression of Maf (also known as c-Maf).
…Overall, our results demonstrate that Malat1 suppression is a hallmark of CD4+ T cell activation and controls IL-10 expression in Th cells. We propose that suppression of Malat1 in activated CD4+ T cells is a critical determinant of optimal immunity to chronic infection.
From 2021, Islam et al discussed the relationship of IL-10 to COVID19 infections27.
Paradoxically, IL-10 can appear to act in a pro-inflammatory manner under certain contexts. IL-10 resistance may be a factor and be related to type 2 diabetes:
…the concurrent elevations in IL-10 and various pro-inflammatory cytokines, and the observed relationship between elevated IL-10 levels and disease severity, suggest that IL-10 is either failing to appropriately suppress inflammation (as observed in other inflammatory conditions (9–11) or acting in a manner that deviates from its traditional role as an anti-inflammatory molecule.Indeed, one explanation for the seemingly paradoxical observation of concurrently elevated IL-10 and pro-inflammatory cytokine levels is the ability of IL-10 to act as a pro-inflammatory and immunostimulatory molecule under certain contexts (6). Another compelling and previously unexplored explanation is the potential escape of activated immune cells from IL-10’s anti-inflammatory action (i.e., IL-10 “resistance”) leading to overexuberant pro-inflammatory cytokine responses. In support of this hypothesis, we have reported resistance to IL-10’s anti-inflammatory action under hyperglycemic conditions in vitro (12, 13) and in individuals with type 2 diabetes (T2D) (12). Importantly, because T2D is a risk factor for increased COVID-19 disease severity and mortality (which is markedly lower with well-controlled blood glucose levels) (14), IL-10 resistance may provide a mechanistic link between hyperglycemia/T2D and adverse COVID-19 outcomes.
And from 2019, and Qu et al performed an in vitro study into how MALAT1 releases epigenetic silencing of HIV-1 replication28.
EZH2: Enhancer of zeste homolog 2, a catalytic subunit that causes viral transcription silencing and maintains HIV-1 latency.
Mechanistically, through an association with chromatin modulator polycomb repressive complex 2 (PRC2), MALAT1 detached the core component enhancer of zeste homolog 2 (EZH2) from binding with HIV-1 LTR promoter, and thus removed PRC2 complex-mediated methylation of histone H3 on lysine 27 (H3K27me3) and relieved epigenetic silencing of HIV-1 transcription. Moreover, the reactivation of HIV-1 stimulated with latency reversal agents (LRAs) induced MALAT1 expression in latently infected cells. Successful combination antiretroviral therapy (cART) was accompanied by significantly diminished MALAT1 expression in patients, suggesting a positive correlation of MALAT1 expression with HIV-1 replication. Our data have identified MALAT1 as a promoter of HIV-1 transcription, and suggested that MALAT1 may be targeted for the development of new therapeutics.
We have above demonstrated that MALAT1 detached EZH2 from HIV-1 LTR and reduces the repressive H3K27me3 of LTR regions. To investigate whether MALAT1-promoted HIV-1 infection was due to the antagonism of EZH2-mediated silencing of viral gene transcription, we further knocked down EZH2 expression with specific shRNA in MALAT1-stably-knocking-out HEK293T cells (Figure (Figure5A and B), and then infected cells with HIV-luc/VSV-G for an additional 24 h. These manipulation of cell gene expression led to a recovery of HIV-1 transcription as quantified by the production of cell-associated HIV-1 gag mRNA (Figure (Figure5C).
Figure 5. MALAT1 antagonizes EZH2-mediated silencing of viral gene transcription to promote HIV-1 replication. (A–C) MALAT1 restores HIV-1 infection by antagonizing EZH2-mediated inhibition. MALAT1-stably-knocking-out HEK293T cells were infected with lentivirus containing EZH2 specific shRNA or off-target controls for 48 h to further knockdown EZH2 expression, cells were then infected with HIV-luc/VSV-G for an additional 24 h. EZH2 knockdown was detected by RT-PCR (A) and western blot (B). Viral infection was detected by quantifying cell-associated HIV-1 gag mRNA (C). (D and E) EZH2 expression in primary CD4+ T cells. Resting CD4+ T cells (1 × 106) were stimulated with or without PHA-P (5 μg/ml) for 3 days, and the endogenous expression of EZH2 was detected by either RT-PCR (D) or western blot (E). (F–H) The double knockdown of MALAT1 and EZH2 rescues HIV-1 infection in primary CD4+ T cells. PHA-P-activated primary CD4+ T cells (1 × 106) were infected with lentiviruses containing MALAT1 or/and EZH2-specific shRNA or off-target controls for 48 h, then cells were infected with replication-competent HIV-1 NL4-3 for an additional 96 h. MALAT1 knockdown was determined by RT-PCR (F); the expression of endogenous EZH2 was detected by RT-PCR and western blot (G); viral replication was quantified by detecting p24gag levels in the supernatants by ELISA (H). Data were presented as mean ± SD. Results were representative of three independent experiments. *P < 0.05 and **P < 0.01 denote significant differences as determined by an unpaired t-test. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6451131/figure/F5/
MALAT1 expression is upregulated in various cancers and thus has been proposed as a prognostic biomarker of metastasis (5,32–37). In HIV-1 infected individuals, we confirmed that successful cART-treatment could significantly reduce expression of MALAT1 (61). Whether MALAT1 could be a biomarker in HIV-1 infection is an interesting topic for future studies.
Taken together, we have identified the critical role of lncRNA MALAT1 in promoting HIV-1 transcription and infection at molecular levels. Our findings may provide a new therapeutic target for combating HIV infection.
Ramirez et al (2022) wrote a literature review on the therapeutic potential of lncRNAs to treat SARS-CoV-2 and endogenous retroviruses such as HIV29.
Of particular interest here, and to read with regard to its association with metastatic cancer, is the suppression of the key tumour suppressor p53 by MALAT1 as well as it correlation to other pathologies including DVT:
Regarding the possible complications of COVID-19 infection, MALAT1 ameliorates deep vein thrombosis (DVT). The mechanism is through inhibition of proliferation and migration of endothelial progenitor cells and finally thrombosis dissolution via the Wnt/β-catenin signaling pathway (102). In addition, MALAT1 reduces the epigenetic silencing of viral transcription by regulating interactions of promoter-enhancer upregulating viral transcription and infection. MALAT1 interacts with STAT1, STAT3, STAT5A, IRF1, and IRF4. MALAT1 is a negative regulator of type I IFN production (10) (Figure 5) and sequesters miR-142-3p and miR-146a-5p to repress their anti-inflammatory activities and its overexpression, thus promoting inflammation (10). Deletion of MALAT1 led to the activation of p53 and its target genes, which affect the normal progression of the cell cycle (15) (Table 2). In addition, in a mouse model of SARS-CoV-2 infection, Malat1 was downregulated by tubulin α1 (TubA1A), the 60S ribosomal protein (Rlp6), and the endoplasmic reticulum protein retention receptor 3 (Kdelr3) (15).
Viruses, cellular lncRNAs and implications for cancer patients
Some viral genes can regulate the levels of cellular lncRNAs to promote viral replication.
Examples of viruses that can interact in this fashion include SARS-CoV, SARS-CoV-2, herpes simplex virus, Marek’s disease virus, HIV, avian leukemia virus and hepatitis B virus (HBV)30. In common with tumours they may also subvert the cell cycle to their own advantage for replication by inhibiting apoptosis31.
Binding of viral proteins to precursors of tumour suppressor p53, the “guardian of the genome” is one way that viruses can inhibit apoptosis32. This can also lead to malignancies such as AIDS-related malignancies from HIV, from human papilloma virus (HPV), from human cytomegalovirus (HCMV) or by spike protein from SARS-CoV-2 virus or synthetic mRNA transfection due to downregulation of p53 and BRCA-133.
P53 suppression can also lead to increased expression of oncogenic KRAS via linked pathways, as explored later.
It is somewhat perplexing that oncologists are still, even now, acting in ignorance to coerce cancer patients into taking experimental gene agents and ineffective influenza vaccines34 in spite of many research papers with findings to the contrary35:
mRNA sourced spike protein inhibits tumour suppressors, as above.
Reinfection after transfection can generate autoantibodies to hundreds of tumour suppressor proteins due to molecular mimicry36.
Lack of long term clinical trials data, nullification of control groups, and an 8 sigma cancer mortality signal above the 5 year baseline37. This should reverse here, the question is by what degree?
If the reason for vaccination is poor immunity then this is an oxymoron as they are less likely to generate an adequate immune response38, or the inflammatory pathways so invoked would be tumorigenic too, such as via IL-6, IL-10, TNFa, TGFb39.
Instead, antivirals that are often not contraindicated but instead are complementary to chemotherapy and anticancer in their own right, such as ivermectin could be administered4041.
Class switching to igG4, with impaired cancer responses haven’t been considered42.
Vaccine/transfection induced lymphopenia due to gp120 binding to DC-SIGN43 and crosstalk to T cell depletion or via ACE2 binding>TGFb upgrading and hematopoietic stem cell depletion44 (cause and effect via pyroptosis45), oncogenic miRNA-21 upregulation46, and so on.
And vaccine induced lncRNA cancer pathways being discussed in this Substack haven’t been adequately explored, let alone verified as endpoints in clinical trials.
Whatever the question, there is only ever one solution, regardless of lack of efficacy, lack of research data, an abysmal safety record and readily available proven alternatives. This needs to stop!
COVID vaccine: Information for people on active cancer treatment
People with cancer* should get vaccinated against COVID-19 unless specifically advised otherwise by their medical team. The Joint Committee on Vaccination and Immunisation (JCVI), the expert group that advises on vaccinations policy, recommends that people on active cancer treatment get vaccinated.
They also make recommendations about the timing of each dose of vaccine in relation to your cancer treatment. Most people in the UK are advised to have two primary doses of vaccine, followed by a booster (a minimum of 3 months later). People whose immune systems are particularly weak – including some people with cancer (specifically those on cancer treatments) – may be advised to have a third primary dose of vaccine (a minimum of 8 weeks after your second dose), followed by a booster (a minimum of 3 months after the third dose).
You will be told by your GP, the NHS Vaccination team or your specialist cancer team if you need a third primary dose. If you had two primary doses before starting immunosuppressive treatment, then you do not need a third primary dose but will still need a booster.
You may be approached about receiving a COVID vaccine at the same time as the flu vaccine. This is acceptable - flu and COVID vaccines are frequently given together.
Your specialist cancer team will be happy to discuss COVID-19 vaccination with you and answer any questions you may have.
If you are going to be receiving drug treatment or radiotherapy to the lung: Ideally you should begin getting vaccinated against COVID-19 before starting your treatment but your cancer treatment can begin while you are waiting for your second dose of vaccine.
Changes to NEAT1 and MALAT1 expression in severe COVID-19 patients
In this paper from 2022, Huang et al performed an analysis of publicly available single-cell datasets with patient severity metrics47.
BAL: “Bronchoalveolar lavage (BAL) is a procedure that is sometimes done during a bronchoscopy. It is also called bronchoalveolar washing. BAL is used to collect a sample from the lungs for testing.48”
BALF: Bronchoalveolar lavage fluid from the above, which is commonly gathered during the diagnostic workup of pulmonary sarcoidosis.
PMBCs: “Human peripheral blood mononuclear cells (PBMCs) are immune cells with a single, round nucleus that originate in bone marrow and are secreted into peripheral circulation. These cells are critical components of the immune system and are involved in both humoral and cell-mediated immunity.49”
rDEGs: “We termed the 50 remaining DEGs recurrent differentially expressed genes (rDEGs) since they were found in multiple cell types and showed differential expression between patients and healthy controls as well as between severities.”
They found that the immunomodulatory lncRNAs NEAT1 and MALAT1 were highly differentially expressed between mild and severe patients in multiple cell types (9) for NEAT1 alone, which was overexpressed in the BAL of severe patients and generally underexpressed in mild patients:
The pro-inflammatory lncRNA NEAT1 passed our rDEG threshold in BAL samples for nine different cell types, more than any other gene in our analysis. These cell types include M1, M2 and intermediate MoMa, NK cells, CD4+ T cells, CD8+ memory T cells, naïve B cells, myeloid dendritic cells, and epithelium/basal cells (Fig 4). NEAT1 is localized to the site of infection and inflammation since it is not differentially expressed in PBMCs. Additionally, among rDEGs, it has one of the highest averages in log2-fold change between severe and mild patients (Fig 4). NEAT1 is overexpressed in BAL of severe patients and underexpressed in mild patients. The epithelial/basal cell group is the exception where mild groups also show NEAT1 overexpression over healthy controls, but expression is still significantly higher in severe patients versus mild patients.
In contrast, MALAT1 showed consistent overexpression in mild patients and underexpression in severe patients:
Another immunomodulatory lncRNA, MALAT1, was the second most frequent rDEG in PBMCs. It passed our rDEG threshold in 6 cell types (tied with ISG15) and 3 cell types in BAL. In BAL derived M1 and M2 MoMa, MALAT1 was underexpressed in mild patients compared to both healthy controls and severe patients. In CD4+ T cells, MALAT1 shows consistent overexpression in mild patients and underexpression in severe patients.In PBMCs, MALAT1 was underexpressed in severe patients versus both healthy controls and mild patients in M1, M2 and intermediate MoMa, NK cells, plasmablasts, and epithelial/basal cells.
NEAT1 overexpression is associated with the activation of inflammasomes and the risk of escalation to highly damaging pyroptosis. If this happens in hematopoietic stem cells it can lead to various long term conditions including lymphopenia and anaemia50 or even impaired immunity of neonates51. I go into more detail on this later.
NEAT1 was the most widely differentially expressed gene across cell types within BAL; it also exhibited a high log-fold change that correlated with disease severity. The ubiquity of NEAT1, its specific localization to BAL cells, and its pro-inflammatory functions suggests that it may be a key mediator of the inflammation seen in severe COVID-19. NEAT1 is a well characterized activator of the NLRP3 inflammasome, as well as the NLRC4 and AIM2 inflammasomes, which in turn amplify the inflammatory response [20]. However, an overactive immune response contributes to lasting tissue damage in severe COVID-19 disease. Intense inflammation through activation of the NLRP3 inflammasome can also lead to pyroptosis, driven by the upregulation of NEAT1 [20, 21]. These highly inflammatory and damaging effects of NEAT1 illustrate how overexpression in severe patients might lead to the inflammatory tissue damage seen in severe COVID-19.
Macrophage skewing to M1 can be induced by overexpression of MALAT1, also mediating further tissue damage. This is somewhat of a paradox as it is also elevating iL-10, as discussed, unless the role of IL-10 has been reversed or neutralised.
M1/M2 describes the two major and opposing activities of macrophages. M1 activity inhibits cell proliferation and causes tissue damage while M2 activity promotes cell proliferation and tissue repair52.
In contrast, overexpression of MALAT1 can lead to a more subdued CD4+ T cell response:
MALAT1 also exerts various immunological effects including the mediation of NLRP3 inflammasome activation [19, 76]. MALAT1 has been linked to M1-like activity in macrophages, promoting inflammation [77]. Our finding that MALAT1 is overexpressed in BAL MoMa of severe versus mild patients suggests that it might be involved in precipitating a shift towards M1 macrophages that exacerbates inflammation. This is further supported by our findings that severe patients show expansion of M1 macrophages and decrease of M2 and intermediate macrophages in BAL, while mild patients show decrease of M1 macrophages. Furthermore, MALAT1 was overexpressed in CD4+ T cells of mild patients. This is also reflective of MALAT1’s protective role in T cells. Loss of MALAT1 expression has been shown to push T cells towards the inflammatory Th1 and Th17 phenotypes while also decreasing Treg differentiation [31]. This function matches our observed increase in abundance of Tregs in mild patients. Thus, the upregulation of MALAT1 in mild patients may be contributing to the more subdued immune response observed in these patients.
Fig 4. lncRNAs NEAT1 and MALAT1 are strongly differentially expressed between severe and mild patients and represent key inflammatory regulators in BAL and PBMC respectively. A. Violin plots showing overall expression level density across patient conditions in the entire dataset. Even at the full dataset scale, these distributions show that NEAT1 is overexpressed in BAL of severe patients while MALAT1 is underexpressed in PBMCs of severe patients. B. Frequency of detection across cell types for rDEGs shows NEAT1 as the most detected rDEG in BAL, with MALAT1 tied for second among rDEGs in PBMC. The top log2-fold change of rDEGs in severe versus mild patients also shows NEAT1 and MALAT1 among the rDEGs with the highest absolute change between severe and mild conditions. C. Visualization of NEAT1 and MALAT1 via UMAP projection shows more cell type localized expression in NEAT1. It is also clearly underexpressed in mild BAL cases. MALAT1 also shows a more subtle but significant underexpression in severe patient PBMCs. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8746747/
We have demonstrated a clear ensemble of differential gene activity associated with severe disease in COVID-19 infection that revolves around the lncRNAs NEAT1 and MALAT1. Their specific activity changes in severe patients, coupled with inflammasome promoting functions, suggest important roles in the COVID-19 hyperinflammatory process. These findings indicate that NEAT1 and MALAT1 may be candidates for treatment targeting or biological marker exploration.
And from 2021, in confirmation of the above, Rodrigues et al evaluated the expression of lncRNAs NEAT1, MALAT1, and MIR3142 in saliva and nasopharyngeal swab from 34 COVID19 positive healthcare workers and 46 who tested negative.
They found a higher expression of NEAT1 and MALAT1 in positive samples than negative. NEAT1 showed higher expression in saliva samples whereas MALAT1 showed upregulation in nasopharyngeal samples. These could be used as biomarkers for infection severity and for the targeting of treatments53.
Positive samples for SARS‐CoV‐2 exhibited a significant upregulation of the lncRNAs NEAT1 and MALAT1 (p < 0.05) (Figure 1). For NEAT1, this difference was observed mainly in saliva samples (p < 0.001), when comparing only NPSs from negative and positive samples, this difference was not observed. In contrast, MALAT1 was observed with higher expression only in NPS positive samples (p < 0.05), when considering only saliva samples, no difference was observed. MIR3142HG expression was detected in less than 17.5% of the samples analyzed and it was removed from further analysis.
Note the log scale on the Y-axis:
FIGURE 1. Relative expression levels of lncRNAs in negative and positive samples for SARS‐CoV‐2. (a) NEAT1 expression including nasopharyngeal swab (NPS) and saliva samples. (b) NEAT1 expression in NPS samples. (c) NEAT1 expression in saliva samples. (d) MALAT1 expression including NPS and saliva samples. (e) MALAT1 expression in NPS samples. (f) MALAT1 expression in saliva samples. *p < 0.05 and **p < 0.01, Mann–Whitney test https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8661855/
Both lncRNAs have been associated with immune system responses. NEAT1 is an inflammatory regulator that promotes activation of inflammasomes in macrophages (Zhang et al., 2019) and induces inflammatory cytokines such as interleukin‐6 (IL‐6) and CXCL8 (Tang et al., 2020). IL‐6 and the NLRP3 inflammasome are primary immune components in responses to SARS‐CoV‐2 infection (Paniri & Akhavan‐Niaki, 2020). MALAT1 also exerts various immunological effects including the mediation of NLRP3 inflammasome activation (Menon & Hua, 2020; Yu et al., 2018). Through NF‐κB and HIF‐1α activation, MALAT1 increases production of inflammatory cytokines, such as IL‐6 and TNF‐α, to promote inflammatory cell infiltration and tissue damage (Tian et al., 2018). Furthermore, MALAT1 has been linked to M1‐like activity in macrophages, promoting pulmonary inflammation and injury (Cui et al., 2019).
And from the previous year, in 2021 Saha et al performed an in silico analysis of the co-regulation of protein coding genes by transcription factor and lncRNAs in SARS-CoV-2 infected cells54. They concluded that differential expression of MALAT1 and NEAT1 was due to the cell type and severity of infection.
This area is poorly understood and they also recommend further studies be conducted.
PBMCs: Human peripheral blood mononuclear cells.
Validation of differentially expressed lncRNA by low throughput assays is absent in most of the studies. In some studies, differences in the expressions of lncRNAs in PBMC and BALF were observed. For example, MALAT1 was decreased in PBMC, while increased in BALF. Similarly, the expression of NEAT1 did not alter in PBMC but was increased in BALF [36]. This difference in PBMC and virus infected cells could be due to the indirect effects of virus infection in PBMC and infected cells. Differential expression of lncRNA in PBMC between severe and mildly affected COVID-19 patients has also been reported. It has been observed that increased level of LINC02207, LINC01127 was associated with the severe COVID-19 group, whereas LINC02084, LINC02446, LINC00861, LINC01871, and ANKRD44-AS1 were associated with the mild COVID-19 group [38]. PVT1 was downregulated in severe COVID-19 patients compared with non-severity [37]. Given thousands of lncRNA coded by the human genome and the lack of consensus of the deregulated lncRNA in SARS-CoV-2 infection, further studies are necessary. Additionally, the mechanism (s) of deregulation of lncRNA, especially by TFs is poorly known.
NEAT1 upregulation is also associated with multidrug resistance and cancer progression via the formation of nuclear paraspeckles.
And both MALAT1 and NEAT1_1 upregulation is associated with repression of tumour suppressor p53.
Nuclear Enriched Abundant Transcript 1 (NEAT1) is a specific structural RNA emerging as a critical component in the progress and development of cancer. NEAT1 forms part of multiple biological pathways, acting through a diverse group of mechanisms. The most important of these is the formation of the paraspeckle, through which it can influence the stability of a tumour to develop resistance to drugs. This review will thus cover the range of effects by which NEAT1 interacts with cancer progression in order to describe the various roles of NEAT1 in chemoresistance, as well as to identify drug targets that protein research alone could not provide.
…Although the full extent of the interactions have not yet been identified, enough research has been carried out in the past few years to begin to address the ways in which paraspeckles are involved in chemoresistance. The current knowledge presents a complex picture in which NEAT1 and even the paraspeckles interact with cancer in different ways. In some cases, these promote (and other times inhibit) cancer growth, resulting in NEAT1 increasing and decreasing chemoresistance in different biological contexts.
Figure 2. Diagrammatic representation of a reconciliation of the current data about the role of NEAT1 in chemoresistance where NEAT1 is activated by numerous factors both tumour promoting and tumour inhibiting. As shown, NEAT1_1 mitigates the oncogenic functions, which is why it is a marker for worse prognosis in colorectal cancer, whilst NEAT1_2 mitigates the tumour suppressor functions. However, it should be noted how both are activated by oncogenic stimulation, which is why paraspeckles tend to form on stimulation of HIF-2, yet its tumour suppressor effect is not enough to balance the oncogenic effect of NEAT1_1. https://pubmed.ncbi.nlm.nih.gov/33143162/
NEAT1 isoforms might have different effects on CSC development with NEAT1_1 aiding in CSC development and NEAT1_2 generally opposing CSC development, such as in the pancreas [37]. NEAT1_1 could therefore also be responsible for gene methylation that induces p53 repression. Therefore, it is possible that NEAT1 exists in a negative feedback loop where p53 induces NEAT1 for paraspeckle formation through NEAT1_2, which aids tumour suppression, whilst NEAT1_1 is also expressed and reduces p53 expression to form the negative feedback loop [19].
CSCs: Cancer stem cells.
Considering the available data, NEAT1 clearly illustrates the importance of lncRNA in cancer and chemoresistance. By being the key structural component to the paraspeckles, as well as through other means, NEAT1 interacts with gene regulatory pathways to bring about changes in gene expression that increase or decrease the ability of tumours to withstand chemotherapy and form CSCs. Thus, the transcript levels of NEAT1 could be a potential biomarker of diagnostic and prognostic value. Despite the fact that a number of the functions of NEAT1 in tumours are unknown, research is beginning to show that the two 3′ processing variants, NEAT1_1 and NEAT1_2, generally have opposing effects in tumours. Even though more studies, which take into account the specific isoform involved in the process investigated are needed to confirm this, the current data is beginning to reveal that by altering the ratio of such 3′ processing variants an effective therapy to decrease the chemoresistance of a tumour may possibly emerge.
DANCR participates in key processes such as stem cell differentiation and tumorigenesis. Xiong et al (2021) conducted in vitro studies and found that it represses apoptosis induction by the chemotherapeutic Doxorubicin through stabilizing MALAT1 expression in colorectal cancer cells56.
And in 2018 Lu et al found through in vitro studies that the oncogene MYC promotes DANCR which in turn promotes cancer by reducing expression of cell cycle inhibitor p21. They found that DANCR is broadly upregulated in many human cancer cell lines and cancers, including most notably in prostate and ovarian cancers57.
Mechanistic investigations indicated that DANCR limited the expression of cell-cycle inhibitor p21 (CDKN1A) and that the inhibitory effects of DANCR loss on cell proliferation could be partially rescued by p21 silencing. In a xenograft model of human ovarian cancer, a nanoparticle-mediated siRNA strategy to target DANCR in vivo was sufficient to strongly inhibit tumor growth. Our observations expand knowledge of how MYC drives cancer cell proliferation by identifying DANCR as a critical lncRNA widely overexpressed in human cancers.
NEAT1 suppression enhances HIV virus production
In 2013, Zhang et al took two T-cell lines, Jurkat and MT4 and infected them with HIV-1 to investigate 83 lncRNAs that have been implicated in diseases ranging from neurodegeneration to cancer in HIV patients58.
They found that NEAT1 is one of several lncRNAs whose expression is changed by HIV infection, and that knockdown of NEAT1 enhances virus production.
The nucleus of higher eukaryotes, such as humans and mice, is compartmentalized into multiple nuclear bodies, an organization that allows for the regulation of complex gene expression pathways that are characteristic of these organisms. Paraspeckles are recently discovered, mammalian-specific nuclear bodies built on a long, non-protein-coding RNA, NEAT1 (nuclear-enriched abundant transcript 1), which assembles various protein components including RNA-binding proteins of the DBHS (Drosophila behavior and human splicing) family.
FIG 2. LncRNA NEAT1 is upregulated in HIV-1-infected cells. (A) Schematic representation of human NEAT1_1 and NEAT1_2. Positions of the sequences amplified by qRT-PCR are indicated with black boxes. (B to E) Jurkat cells (B), MT4 cells (C), THP1 cells (D), or THP1 cells differentiated with PMA (E) were infected with VSV-G-pseudotyped HIV-1 NL4-3. Three days after infection, NEAT1 expression levels were analyzed by qRT-PCR. Results from three separate experiments are shown as mean values ± SD. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3560530/
FIG 3. Knockdown of NEAT1 decreases paraspeckles. HeLa cells were transfected with 50 nM control siRNA or NEAT1 siRNA. (A) At 2 days posttransfection, the knockdown efficiency was confirmed by qRT-PCR. Results from three separate experiments are shown as the mean values ± SD. (B) Twenty-four hours after transfection of control (top) or NEAT1 (bottom) siRNA, HeLa cells were retransfected with a GFP-PSF-expressing plasmid for 24 h. The cells were then visualized by confocal microscopy for GFP-PSF (green), endogenous PSP1 stained with specific antibody (red), and nuclei stained with 4′,6-diamidino-2-phenylindole (DAPI) (blue). The number of paraspeckles per cell was quantified by visual counting of more than 50 cells. Results are shown as mean values ± SD. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3560530/
Here, we report the first evidence of NEAT1 as an lncRNA involved in HIV-1 replication. We show that HIV-1 infection increases NEAT1 RNA expression by 5- to 10-fold over that for uninfected cells (Fig. 2). Using two different approaches, siRNA and antisense DNA, we demonstrated that the knockdown of NEAT1 enhanced virus production by increasing nucleus-to-cytoplasm export of Rev-dependent INS-containing HIV-1 transcripts.
NEAT1 induced paraspeckle bodies can act as a storage pool for unspliced HIV-1 RNA’s, diverting them away from being spliced as a prelude to viral replication via the nucleolus. Many viruses use this to complete several essential steps of the viral cycle59:
That the knockdown of NEAT1 reduces paraspeckle bodies and results in increased cytoplasmic expression of HIV-1 INS-containing mRNAs suggests that paraspeckles may indeed represent the long-postulated nuclear compartment for storing HIV-1 Rev-dependent INS-containing RNAs that are diverted away from splicing (65). We consider that the nuclear pool of unspliced HIV-1 RNAs may contain two subpools, one destined for splicing/degradation and the other destined for storage in nuclear paraspeckles. A small change in the latter pool from downregulation of paraspeckles could have a significant effect on cytoplasmic unspliced RNA levels without dramatically changing overall nuclear unspliced RNA levels.
…For cellular transcripts, a paraspeckle depot may be a means for stabilizing and maintaining RNAs that might otherwise be degraded. This type of RNA storage followed by release (when required) offers a more rapid and resource-efficient way than de novo RNA biosynthesis for providing needed RNA for rapid use in responding to stress. Thus, HIV-1 seemingly has co-opted this cellular mechanism to also store its excess unspliced INS-containing viral RNAs in paraspeckle bodies.
Also from 2021, it was not known if SARS-CoV-2 also induced paraspeckle formation, but it was very likely as evidenced by upregulation of NEAT160. Could these also act as RNA reservoirs for the virus and/or transfected mRNA? You would think it useful to know these things before administering such agents en masse?
ISGs: IFN-stimulated genes.
IAV: Influenza virus.
HTNV: Hantaan orthohantavirus.
It is unknown whether paraspeckles is induced in response to SARS-CoV-2 infection; although expression of NEAT1, the indispensible component of paraspeckles was increased in our analysis as well as various viral infections including IAV, HIV and HTNV. Thus similar mechanism might be operative in SARS-CoV-2 infected cells to increase ISGs.
NEAT1 suppression enhances Hantaan virus production
Further to the above, from 2021 and Yang et al performed an in vitro analysis of Hantaan infected mouse bone marrow-derived macrophages and found that silencing the NEAT1-2 isoform hinders inflammatory macrophage activation and facilitates HTNV propagation, whereas in contrast enhanced NEAT1-2 transcription effectively restrains viral replication61:
Hantaan virus (HTNV) is prevalent in Asia and is the leading causative agent of severe hemorrhagic fever with renal syndrome (HFRS), which has profound morbidity and mortality.
In this study, we examined the gene expression profile of macrophages at different time points after HTNV infection through high-throughput RNA sequencing (RNA-seq) and observed elevated NEAT1 expression, as well as the enhanced cholesterol synthesis process, at the early stage. Interfering with NEAT1-2 transcription could restrain inflammatory macrophage activation and thus facilitate HTNV propagation in a macrophage-endovascular coculture system, while forced expression of NEAT1-2 stimulated macrophages to secrete various cytokines and chemokines, such as TNFα and IFNα, hence restricting HTNV infection. Mechanistically, we found that NEAT1-2 might upregulate Srebf2 expression, bind to SREBP2, and positively influence its proinflammatory activity. Moreover, the NEAT1-2 transcription level in monocytes was associated with viral load and disease severity. These results reveal a previously unrecognized role of NEAT1 in metabolism and macrophage polarization and suggest that the altered expression of NEAT1-2 might be a potential biomarker for predicting HFRS progression.
Its not that easy to see here, but si-NEAT1-2 (the silenced form) promoted macrophage polarisation (“skewing”) to M2 away from the inflammatory, antiviral M1 types:
FIGURE 2. Enhanced M1 Polarization by NEAT1-2 to Constrain HTNV Propagation.(A) (I) Schematic diagram of the coculture system. (II) RNAi efficiency of NEAT1-2 silencing in hMDMs confirmed by qRT-PCR. (III) RNAi efficiency of NEAT1-2 in hMDMs confirmed by RNAScope. NC, negative control with scrambled RNAs. (B) (I) qRT-PCR analysis of M1-related genes in hMDMs from the coculture system shown in 2A-(I). (II) qRT-PCR analysis of M2-related genes in hMDMs from FIGURE 2 | the coculture system. (C) Immunoblot analysis of HTNV NPs in hMDMs from the coculture system. (D) qRT-PCR analysis of HTNV S segment hMDMs from the coculture system. (E) Enzyme-linked immunosorbent assay (ELISA) detection of IL-8 and CCL5 concentrations in the bottom medium in the coculture system. (F) Immunoblot analysis of the indicated proteins in HUVECs in the coculture system. (G) Viral titers of HUVECs assessed by the improved enzyme-linked focus formation assays. (H) (I) Representative flow cytometry data for TNFα and IL-10 production in NEAT1- or NEAT1-2-overexpressing hMDMs at 12 hpi (MOI = 5). (II) Statistical analysis of data from (I). (I)(I) Representative flow cytometry data for HTNV NP expression in NEAT1- or NEAT1-2-overexpressing hMDMs at 36 hpi (MOI = 5). (II) Statistical analysis of data from (I). (J) ROS measurement of hMDMs from I-(I). Data are shown as the mean ± SEM and are representative of three independent experiments. Each point represents a single sample (n = 4 in each group). Analysis was performed using the unpaired Student’s t-test (A–G) or one-way ANOVA (H–J). *p < 0.05, **p < 0.01, and ***p < 0.001. https://pubmed.ncbi.nlm.nih.gov/35495674/
In conclusion, we found in this study that lncRNA NEAT1-2 regulated HTNV-induced M1 polarization, constraining viral propagation and spread between cells. NEAT1-2 not only promoted SREB2 expression and cholesterol synthesis at the early infection phase but also interacted with SREBP2 and strengthened SREBP2-mediated inflammatory responses. Moreover, NEAT1-2 expression in monocytes was shown to be negatively correlated with HFRS progression. These results revealed a novel biomarker for the evaluation of host immune functions against HTNV infection and suggested a potential therapeutic target for the clinical treatment of HFRS.
DANCR and NEAT1 suppression is associated with enhanced COVID19 inflammatory pathobiology
In 2020, Meydan et al selected inflammation-prone men and women of diverse ages among the cohort of Genome Tissue expression (GTEx) and mined RNA-seq datasets from their lung, and blood tissues62.
They then performed bioinformatics-based network analyses and performed statistical analysis of results from brain cell culture and infection tests with SARS-CoV-2 and H1N1 viruses.
DANCR appears to act in much the same way as NEAT1 and was also suppressed in severe infections:
In lung tissues from 57 inflammation-prone, but not other GTEx donors, we discovered sharp declines of the lung pathology-associated ncRNA DANCR and the nuclear paraspeckles forming neuroprotective ncRNA NEAT1. Accompanying increases in the acetylcholine-regulating transcripts capable of controlling inflammation co-appeared in SARS-CoV-2 infected but not H1N1 influenza infected lung cells. The lung cells-characteristic DANCR and NEAT1 association with inflammation-controlling transcripts could not be observed in blood cells, weakened with age and presented sex-dependent links in GTEx lung RNA-seq dataset. Supporting active involvement in the inflammatory risks accompanying COVID-19, DANCR’s decline associated with decrease of the COVID-19-related cellular transcript ACE2 and with sex-related increases in coding transcripts potentiating acetylcholine signaling. Furthermore, transcription factors (TFs) in lung, brain and cultured infected cells created networks with the candidate transcripts, indicating tissue-specific expression patterns. Supporting links of post-infection inflammatory and cognitive damages with cholinergic mal-functioning, man and woman-originated cultured cholinergic neurons presented differentiation-related increases of DANCR and NEAT1 targeting microRNAs. Briefly, changes in ncRNAs and TFs from inflammation-prone human lung tissues, SARS-CoV-2-infected lung cells and man and woman-derived differentiated cholinergic neurons reflected the inflammatory pathobiology related to COVID-19.
Graphical abstract: Study workflow - DANCR and NEAT1 interact with inflammatory markers in a sex- and tissue-specific manner(A) Segregating blood and lung samples from the GTEx dataset of healthy post-mortem donors into resilient and inflammation prone. Samples expressing high levels of seven of the inflammation biomarkers IL-1B, IL-6, NFKB1\2, REL, RELA\B were defined as inflammation-prone, and others as resilient. Differential expression analysis highlighted DANCR as a lncRNA DE between the two groups. Parallel analysis of cultured lung epithelial cells infected with SARS-CoV-2 found DANCR’s decline in infected cells. (B) Both GTEx post-mortem men and women lung and brain tissues and lung-derived cell culture infected with SARS-CoV-2 showed DANCR and NEAT1 changes in inflammatory biomarkers, part of which show modified levels under COVID-19 infection. Three different TF controllers of DANCR and NEAT1 can mediate the inflammatory tone in diverse organs (SPI1 (blue) in cortex, RUNX3 (yellow) in lung and TTF2 (green) in lung epithelial cells). DANCR and NEAT1 can block inflammation via interacting with other ncRNAs, sponging miRs, or affecting TFs like STAT3. Red, flat-headed and green arrows indicate expression blockade and induction of expression, respectively. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7581732/
In particular, we noted DANCR, which showed a pronounced decline in the GTEx inflammation-prone lung tissues (p<0.1e-13, t.test). The 855 nucleotides long DANCR transcript is produced from chromosome 4q12 (59), and is known to modulate the action of CTNNB1 (catenin beta-1), which is actively involved in respiratory infections and sepsis (60–62). Previous studies show that DANCR can trigger the pro-inflammatory STAT3 activation via the IL-11-JAK2 pathway (63). STAT3, in turn, activates IL-1b, IL-6, NFkB1, REL, and RELB (64–66), as well as the transcription factor (TF) SPI-1 (Pu.1) which further interacts with TNF, NFkB2, RELA, DANCR, and the NEAT1-associated p54nrb paraspeckle protein (67–71).
…Both DANCR and the sepsis-related neuroprotective lncRNA NEAT1, which initiates and maintains the membrane-less organelle of nuclear paraspeckles (30, 33, 39), were suppressed in SARS-CoV-2 infected cells (Figure 2C) (see corresponding Resource in Supplementary Table 1 for the full list of lncRNAs changed in NHBE and A549 cells infected with SARS-CoV-2, in NHBE cells infected with pandemic H1N1, and in lung inflammatory samples). Together, these observations indicated that inter-related changes in DANCR, NEAT1, and NFkB1 might play specific regulatory roles in the natural history of SARS-CoV-2 infection.
DANCR and NEAT1 suppression is also associated with cognitive impairment, aka “brain fog”, worsening with age, and due to inflammation mediated in part by consequential suppression of acetylcholine expression:
The inflammatory association of the DANCR- and NEAT1-related networks and the inverse changes of DANCR and AChE along age predicted body-brain messages affecting the cholinergic network. Compatible with recent reports of cognitive damages in infected patients (23, 26), we considered that these interactions may reflect a response to the infection-induced inflammation, which might mitigate neuronal functioning and damage cognition during and after the acute phase.
…Intriguingly, differentiated cholinergic neurons presented a selective increase of one of the splice variants of DANCR, DANCR-205 (748 bp; p<0.063), accompanied by upregulation of the nuclear paraspeckles-forming neuroprotective lncRNA NEAT1 (Figure 3G; p<0.09). Thus, both lung cells and brain neurons, but not blood cells, show informative changes in DANCR and its related inflammatory transcripts. Cholinergic neurons showed concomitant increases of expression of DANCR and NEAT1 with ChAT levels reflecting their acetylcholine producing capacities. As acetylcholine binding to the nicotinic acetylcholine receptor alpha7 can block NFkB and inflammation, we propose that DANCR and NEAT1 elevations may modify the inflammatory status of neurons.
Several microRNA’s are differentially expressed (DE) due to infection, of which some of these help suppress DANCR & NEAT1 expression. The interaction of oncogenic miR-22 is of interest here, as are miR-19a-3p and miR-335-5p, and the male/female related differences. These lncRNAs appear to be acting as miRNA sponges.
I always like seeing the word “strikingly” in a paper!
Strikingly, out of 32 differentially expressed (DE) miRs under SARS-CoV-2 infection, 21 were also DE in cholinergic neurons of either male or female lines following cholinergic differentiation (or both; Figures 6A, B). Of these miRs, particularly interesting are the pain-regulating miR-21-5p (103) and miR-22-3p, the levels of which presented opposite directions of change under cholinergic differentiation and SARS-CoV-2 infection, with male-originated cells showing a drastically stronger effect than female-originated ones (Figure 6C). Compatible with the ‘sponging’ property of both DANCR and NEAT1, the infection-related DE miRs showed intriguing interactions with DANCR, NEAT1, and the selected set of inflammation-related transcripts, eleven of which are targeted by 16 out of these 32 DE miRs; furthermore, both NEAT1 and DANCR carry complementary binding sites to 24 out of these 32 DE miRs, and 18 of the miRs targeted by NEAT1 and DANCR also target at least one of the tested inflammatory transcripts (Figure 6D). Specifically, miR-19a-3p and miR-335-5p target both NEAT1 and DANCR as well as couple of inflammation controlling transcripts and are downregulated under SARS-CoV-2 infection. Thus, the changes in DANCR and NEAT1 may lead to the infection-related impact of those miRs and regulate the inflammation signature associated with them in epithelial cells, albeit in a sex-related manner.
Anti-cholinergic medications may be contraindicated for some patients as a consequence of these findings:
We discovered the lung pathology-associated lncRNA DANCR and the nuclear paraspeckles forming neuroprotective lncRNA NEAT1 as potentially involved in the susceptibility to and consequences of COVID-19, in conjunction with acetylcholine and inflammation-regulating transcripts. Notably, these observations were undetectable in blood samples, weakened with age and presented sex-dependent links in cholinergic neurons, highlighting lung cells as a preferable site for COVID-19 diagnostics. At the same token, these findings call for greater appreciation of the impact of medications with anti-cholinergic effects on post-infectious cognitive sequelae; these agents are commonly used as supportive measures in intubated and spontaneously breathing hospitalized patients. Taken together, our findings may assist future management of COVID-19 patients, as they may exhibit diverse expression profiles of molecular regulators such as DANCR and NEAT1, in conjunction with previously depicted inflammatory mediators such as IL-1, IL-6, TNF, and more.
Recent COVID-19 reports raise the importance of neurological complications inherent in the disease course (24, 134), indicating causal involvement of cholinergic-associated ncRNAs in CNS pathology (e.g. the AChE-targeting and inflammation-regulating miR-132 (45)). Furthermore, aged patients under prolonged anti-cholinergic medications are at increased risk of cognitive decline (135), and our findings of the apparent cholinergic-DANCR-NEAT1 links raise the question if such patients would also display excessive susceptibility to failed ncRNA response and hence harsh cognitive consequences of COVID-19.These effects may be augmented by adverse mental derangement associated with intensive-care settings and mechanical ventilation, but the relatively common loss of smell and taste in reports so far (22) suggests that SARS-CoV-2 has inherent neurological effects.
From a clinical pearls article on anticholinergic medications) by Ghossein et al (2022)63:
Anticholinergic medications are used to manage and treat a wide range of diseases. This activity illustrates the indications, action, and contraindications for anticholinergic drugs as valuable agents in managing cholinergic toxicity, urinary incontinence, Parkinson disease, respiratory disorders, cardiovascular disease, and numerous other diseases.
Ipratropium and tiotropium: Function to dilate the bronchi and relieve shortness of breath and FDA approved for the use in patients with chronic obstructive pulmonary disease.[6]
Benztropine and trihexyphenidyl: Used to counter reduced dopamine levels and relieve symptoms of Parkinson disease [6]
Oxybutynin and tolterodine: FDA approved for the treatment of urge incontinence and detrusor hyperactivity[6]
Oxybutynin has an off-label use for the treatment of hyperhidrosis.
Diphenhydramine and other anti-histamines: FDA approved as a sleeping aid
Scopolamine: Used as a prophylactic anti-emetic[8]
Atropine: Used to dilate pupil during retina visualization and in the treatment of cholinergic toxicity[9]
Vecuronium and Succinylcholine: Antinicotinic medications used as a neuromuscular blockade in surgeries[6]
Mecamylamine: Used strictly in research settings as a ganglionic blocker[6]
Glycopyrrolate: A type of quaternary amine used in anesthesia to decrease salivary and tracheal secretions[10]
Medications like antipsychotics, tricyclic antidepressants, and diphenhydramine (when used to treat allergies) possess anticholinergic properties despite not being responsible for their therapeutic qualities.
And most importantly to see contraindicated for patients on vents:
The medication most commonly associated with high anticholinergic burden in our cohort was midazolam, one of several benzodiazepines judged to have anticholinergic properties by the ADS (2, 24). The properties of benzodiazepines have been long discussed in the context of anticholinergic activity.64
SARS-CoV-2, Cardiovascular Diseases, and Noncoding RNAs
In this paper from 2021 by Natarelli et al, the connected triad of these three factors is discussed65.
NEAT1 expression is disrupted by a wide range of viruses other than SARS-CoV-2, including hepatitis C & B, HIV, in encephalitis and by influenza infections :
On the other hand, studies on the role of lncRNAs in viral infections are still scarce. Few lncRNAs have been reported to promote or inhibit viral replication upon infection. NEAT1 [62], EGOT [63], NRON [62], and lncRNA-CMPK2 [63] have been reported as lncRNAs regulated by the hepatitis C and B viruses (HCV and HBV) while NEAT1 is upregulated in HIV, encephalitis, and influenza infections [62,63].
Even though a detailed description of ncRNA synthesis and delivery is out of the scope of this review, it is worth mentioning that ncRNA can interact with viral genomes either inside the expressing cells, and in particular, within target cells, after being delivered to in the blood vessels. Circulating ncRNAs may originate either from immune cells or from endothelial cells of other organs. In the blood stream, miRNAs circulate as microparticles cargo (exosomes, microvesicles, and apoptotic body) or are associated with specific RNA-bind proteins, such as Argonaute-2 (Ago2), and with lipoproteins [41].
Viral lncRNAs can interfere with or be incorporated into the host cell genome and disrupt miRNA expression:
Interestingly, recent data reported the existence of viral lncRNAs, named virus-encoded lncRNAs and chimeric lncRNAs, which can interfere with or are incorporated in the host cell genome. Among these are EBER1/2, Adenovirus (AdV) virus-associated RNA I and II (VAI, VAII RNA), and sfRNAs [64]. Adenovirus (AdV) virus-associated RNA (VA RNA) is a human essential pro-viral ncRNA known to relieve the cellular antiviral blockade of protein synthesis. Recent findings revealed that VA RNAs interfere with the Dicer-mediated miRNAs gene silencing [65]. In detail, VA RNA fragments can inhibit the export of miRNA precursors in the cytoplasm by saturating the nuclear export protein Exoprtin 5 and can saturate the ribonuclease Dicer to interfere with miRNA biogenesis. Additionally, VA RNA competing fragments can be processed by Dicer and incorporated into the RISC complex as “mivaRNAs”, thus inhibiting the mediators of the immune system [65]. Hence, viral lncRNAs may represent a novel attractive alternative in designing antiviral therapies.
Other investigations indicated miR-21 as the miRNA with the best binding to the SARS-CoV-2 genome [74]. MiR-21 regulates cardiac structure and function by modulating the ERK–MAP kinase signaling pathway in cardiac fibroblast, and it is increased during heart failure, where acts as a promoter of interstitial fibrosis and cardiac dysfunction [75]. Human miR-23b and miR-126 have also been identified as potentially able to bind SARS-CoV-2 genes [59]. Notably, miR-126-3p and miR-126-5p are known regulators of endothelial vascular biology and leukocyte adhesion [76], and their expressions are significantly affected in patients with CVD [35,77]. Recently, the work of Dongchao Lu and colleagues supported the role of miR-200c in binding and inhibiting the ACE2 transcript in rat and human iPSC-derived cardiomyocytes [78]. Since miR-200c is upregulated in CVDs, controversy still exists on the effects related to ACE2 in CVD and the role of miR-200c in COVID-19 infection.
As for miRNAs, lncRNAs and circRNAs are known to be involved in several physiological cell functions, including immune tolerance. Observational studies indicate that lncRNA H19, which promotes pulmonary arterial hypertension (PAH), promotes abdominal aortic aneurysm in mice and pigs. LncRNA LIPCAR emerged as a plasma molecular biomarker of acute myocardial infarction (AMI), since its levels increased during cardiac remodeling progression [79] concomitantly with increased levels of ACE2 proteins [80]. However, data demonstrating a direct involvement of lncRNAs and circRNAs as a molecular link between CVD, MI, and SARS-CoV-2 infection are missing.The data reported are still indirect and putative but potentially support the role of human lncRNAs as novel targets in designing antiviral therapies against viral infection in CVD patients. Among these, studies on bronchial epithelial cells and lung tissues from patients infected by SARS-CoV-2 reveal that certain host-derived lncRNAs and circRNAs, such as MALAT1, NEAT1, and HRCR, are affected in patients infected by COVID-19 [79]. Additionally, LIPCAR, H19, ANRIL, MIAT1, and SENCR were modulated in patients with CVD [79,81].
The authors were later proven correct, unfortunately, in this concluding statement from their 2021 paper:
Although the efficacy of RNA-based novel vaccines is higher than that of previous vaccines, the high mutation rate reported in the Spike protein suggests that therapies based on Spike protein may be ineffective in a long-term perspective.Moreover, unlike many conventional vaccines, the use of Spike to activate the host antiviral response would inevitably require the constant production of new vaccines able to protect against new SARS-CoV-2 variants. This procedure could not be favorable in terms of cost–benefits and would lead patients with cardiovascular problems to undergo vaccination cycles not devoid of possible side effects.Are we therefore considering the wrong target in designing RNA-based vaccines?
COVID19, synthetic mRNA gene agents and lncRNA interactions
This paper by Moazzam-Jazi et al from 2021 posed the following hypothesis66:
In contrast to the 3′ part of the SARS‐CoV‐2 genome, the 5′ part can interact with many human lncRNAs. Therefore, the mRNA‐based vaccines will not show any side effects because of the off‐label interactions with the human lncRNAs.
…The human lncRNA‐virus genome interaction sites appear to restrict the ORF1ab gene and rarely span NSP5 or NSP6. Consistent with our findings, Vandelli et al demonstrated the 5′ end of the viral genome is highly structured and can interact with various human proteins. 15 Also,the viral portion of SARS‐CoV‐2 harbouring the sequence coding spike protein tends to interact neither with human proteins nor with human lncRNAs, implying that the mRNA‐based vaccines will not show the possible side effects because of the off‐label interactions with these macromolecules.
So that’s all good then, as you were.
No conflict of interest declaration was made.
LncRNA H19 upregulation
We have already reviewed several paper showing that SARS-CoV-2 does interact, and there is in fact evidence that spike protein mRNAs are indeed interacting too, both from the virus and also from synthetic experimental mRNA’s, but there are key differences, especially with interferon mediated antiviral responses.
From 2021, and Natarelli et al conducted in silico analysis of the SARS‐CoV‐2 Wuhan-Hu-1 dataset and then, after further interaction analysis, used the RNAfold web tool to predict the interaction propensity between lncRNAs and the Spike transcript, SARS-CoV-2 5′, or 3′UTR67.
Here, we report a computational study demonstrating the existence of target motifs in the SARS-CoV-2 genome suitable for specific binding with endogenous human micro and long non-coding RNAs (miRNAs and lncRNAs, respectively), which can, therefore, be considered a conceptual background for the development of miRNA-based drugs against COVID-19. The SARS-CoV-2 genome contains three motifs in the 5′UTR leader sequence recognized by selective nucleotides within the seed sequence of specific human miRNAs. The seed of 57 microRNAs contained a “GGG” motif that promoted leader sequence-recognition, primarily through offset-6mer sites able to promote microRNAs noncanonical binding to viral RNA. Similarly, lncRNA H19 binds to the 5′UTR of the viral genome and, more specifically, to the transcript of the viral gene Spike, which has a pivotal role in viral infection. Notably, some of the non-coding RNAs identified in our study as candidates for inhibiting SARS-CoV-2 gene expression have already been proposed against diverse viral infections, pulmonary arterial hypertension, and related diseases.
…We considered an MFE threshold of <−20 for miRNA binding sites (BS) predicted at the viral 3′UTR or at the Spike mRNA transcript (3′UTR).
…The interaction propensity between lncRNAs and the Spike transcript, SARS-CoV-2 5′, or 3′UTR was predicted using the RNAfold web tool [24], following the prediction of lncRNA putative secondary structures using the RNAfold web tool. Colors represent base pair probabilities.
…Similar to miRNAs, lncRNAs can interact with RNA, DNA, and proteins, and form RNA–RNA, RNA–DNA, and RNA–protein complexes, leading to the regulation of gene expression via multiple mechanisms, including modulation of transcription, mRNA stability, and translation.
The strongest lncRNA to spike mRNA interaction was with H19:
Figure 5. Interaction propensity of lncRNAs involved in pulmonary arterial hypertension, antiviral response, and inflammatory diseases. (a) Heat map of lncRNA interaction propensity with SARS-CoV-2 5′UTR, 3′UTR, and with Spike mRNA using IntaRNA, RNAup, and RNAplex. Significantly binding sites for Spike were identified for cytoplasmic (b) lncRNAs H19 (and for 5′UTR), (c) FENDRR, and (d) LINC01505. Nuclear lncRNAs were used as negative control. Rectangles represent the zoomed view of lncRNA interaction loops. LncRNA minimum free energy secondary structures were predicted using RNAfold web tool. Colors represent base pair probabilities. * p < 0.05; ** p < 0.01; *** p < 0.001. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7931055/
Not only is H19 binding with Spike mRNA, it appears to be more pathogenic because of this:
Three LncRNAs, such as FENDRR, HOTAIR, and LINC01505, were found to potentially interact with Spike mRNA. Given that the secondary structure of HOTAIR has been experimentally determined [72], we were able to map its interacting regions with the Spike mRNA in Domain 2, elements H16–H21, and in Domain 4, elements H50–52. LINC01505 also showed a binding propensity for SARS-CoV-2 3′UTR (Figure 5c,d). Notably, H19 has been found to promote the pathogenesis of pulmonary arterial hypertension (PAH) [73], suggesting that it might also contribute to SARS-CoV-2 acute pulmonary injury.
…In this work, we consider lncRNAs as well, which can interplay with miRNAs, for example by acting as sponges of miRNAs, and as we demonstrated in this work, can bind to the viral genome and Spike transcript.
H19 was also found to be dramatically upregulated in BALF (bronchoalveolar lavage fluid) of COVID-19 patients, consistent with higher viral loads68.
Interactions with spike protein mRNA appears to be a contributory factor, as is suppression of p53 by multiple pathways:
Spike protein localizes in the nucleus and inhibits DNA damage repair by impeding key DNA repair protein BRCA1 and 53BP1 recruitment, as discussed earlier.
MALAT1 deactivates p53, as discussed earlier.
Spike protein mRNA derived MiR-21 suppresses BMAL1 which in turn inhibits gene promoter activity for p53 expression69.
Suppression of p53 then induces another pro-tumour pathway via H1970: KRAS.
In a previous work, we reported a negative regulation of H19 promoter activity by the tumor suppressor p53 that would be mediated by protein/protein interactions.
To expand on this, upregulated H19 acts as a molecular sponge to inhibit the tumour suppressor let-7 family of miRNAs. Thus regarding cancer risk, less H19 is better:
Abundantly expressed in fetal tissues and adult muscle, the developmentally regulated H19 long noncoding RNA (lncRNA) has been implicated in human genetic disorders and cancer. However, how H19 acts to regulate gene function has remained enigmatic, despite the recent implication of its encoded miR-675 in limiting placental growth. We noted that vertebrate H19 harbors both canonical and noncanonical binding sites for the let-7 family of microRNAs, which plays important roles in development, cancer, and metabolism. Using H19 knockdown and overexpression, combined with in vivo crosslinking and genome-wide transcriptome analysis, we demonstrate that H19 modulates let-7 availability by acting as a molecular sponge. The physiological significance of this interaction is highlighted in cultures in which H19 depletion causes precocious muscle differentiation, a phenotype recapitulated by let-7 overexpression. Our results reveal an unexpected mode of action of H19 and identify this lncRNA as an important regulator of the major let-7 family of microRNAs.
TMPRSS2 expression can also be induced by H19 acting as a miRNA sponge.
SARS-CoV-2 can then use this as a means to infect cells. In other words unintended vaccinal promotion of H19 can paradoxically lead to increased SARS-CoV-2 viral loads.
To deepen further our analysis, we have to consider another type of regulation of gene expression related to long non‐coding RNA (LncRNA), which are non‐coding RNAs capable to act as sponges for miRNAs and coding mRNAs. We have identified the lncRNA H19, harbouring canonical and non‐canonical binding sites for the let‐7 family of miRNAs. In vitro and in vivo analysis demonstrated that H19 could modulate let‐7 availability. Among many oncogenic roles, H19 is able to modulate the expression of IL‐6 via sponging the let‐7 family members in turn possibly inducing TMPRSS2 expression by sequestering its target miRNAs.We could hypothesize that H19 action may be likely responsible for a greater virus ability to infect cancer cells.
A downstream consequence of let-7 sponging is upregulation of oncogenic K-RAS protein. Mutated forms are especially oncogenic.
Hat tip to Genervter for raising awareness of this.
Ibrahim et al (2020)73 discuss the role of KRAS in colorectal cancer:
Colorectal cancer (CRC) is the third most commonly diagnosed cancer and the fourth leading cause of cancer death worldwide. 1 CRC is a multistep carcinogenesis caused by the accumulation of genetic mutations and alteration in signaling pathways. 35% of CRC is due to gene mutations,2 with KRAS gene mutations accounting for 40% of these CRC cases and NRAS accounting for nearly 5%.3
…RAS proteins are GTPases that regulate the RAS signaling pathway that control cell proliferation and cell survival and are often mutated in human cancers. Human RAS genes are comprised of Kirsten RAS (KRAS), Neuroblastoma RAS (NRAS) and Harvey RAS. The first 85 amino acid residues at the N-termini of all three ras isoforms are highly related - possessing a role in GTPase activity.5 However, different isoforms of ras protein are involved in different types of cancer. For example, KRAS mutations are frequently found in solid tumors such as lung, colorectal and pancreatic cancers,6 whereas NRAS are found mostly in hematopoietic tumors and melanomas.6,7
85% of KRAS gene mutations occurs in codons 12 and 13 of exon 2, while the remaining 15% is found within codon 61 of exon 3.8During carcinogenesis, activation of KRAS proteins was not required for tumor initiation, however the activation significantly increased tumor incidence and accelerates tumor growth.9KRAS mutations have been detected in both early and late CRC, indicating that KRAS mutations might occur in the early stage of tumor development.6
Let-7 family
Lethal-7, or mostly termed as let-7, is one of the earliest discovered miRNAs. Let-7 is negatively regulated with KRAS expression in CRC.26An overexpression of let-7 reduces KRAS and DNA damage repair genes, such as RAD51 and CDC25.27 There are currently thirteen known members in the let-7 family, locating at nine different loci. All the let-7 family members have highly similar sequence and share a seed region (GAGGUAG), which is a nucleotide motif, an important component for RNA-induced silencing complex target recognition.27
Of the 13 members, Let-7a-1-5p is most frequently downregulated in CRC. Administration of let-7a-1-5p precursor demonstrated a suppressive effect on growth and proliferation in human colon cancer cells, DLD-1 and SW480 cell lines.26 Overexpression of let-7a-1-5p reduces KRAS and c-myc protein expression, but not the KRAS and c-MYC mRNAs.26 Overexpression of let-7a decreases the radiosensitization of cells during therapy.27
Although let-7 is one of the first miRNAs discovered, the functional roles of let-7 family members have yet to be understood. Choo et al. demonstrated that let-7d-3p/5p are both co-expressed in colon cancers - Let-7d-3p specifically downregulates KRAS whereas let-7d-5p upregulates IGF1R and THBS1.11 In a study of 49 Stage II CRC patients, the upregulation of let-7b and let-7d were associated with microsatellite stability (MSS).28Let-7d-3p targets KRAS protein in vascular smooth muscle cells and a transfection of let-7d-3p decreases the KRAS protein - cell growth were reduced, and the G1 cell cycle was induced when compared to the G2/M phase.29A recent study by Gunel et al. showed that let-7d- 3p downregulates KRAS and HMGA2 in epithelial ovarian cancers.30 Further study on let-7d and its involvement in the cell cycle is required.
There are multiple let-7 complementary sites (LSCs) within the 3'UTR of KRAS mRNA.
T to G base substitution on rs712 within the 3’UTR of KRAS mRNA weakens the binding to let-7, increasing the expression of constitutively active KRAS and thus activating the RAS-MAPK pathway which results in carcinogenesis.
H19 also regulates smooth muscle cell functions and upregulation is associated with an increased risk of aortic rupture/dissection (AD) through sponging miR-193b-3p.
From 2021, and Ren et al conducted an vitro analysis using aortic tissue samples from AD sufferers and in vivo analysis using mice that had been artificially induced to be susceptible to AD by implanting a micropump to dose them with Ang II (angiotensin II)74:
Results: LncRNA H19 was abnormally high-expressed in thoracic aorta tissues of AD patients, and it could competitively bind to and inhibit miR-193b-3p. In the PDGF-BB group, the expressions of H19, matrix metallopeptidase (MMP) 2 (MMP-2) and MMP-9 were up-regulated and the expressions of miR-193b-3p, α-SMA, and SM22α were down-regulated; moreover, the proliferation and migration rate of HASMCs were increased. However, H19 silencing reversed the regulation of PDGF-BB on HASMCs. More interestingly, miR-193b-3p inhibitor could partially reverse the effect of H19 silencing. In addition, the above results were verified by animal experiments, showing that shH19 and up-regulated miR-193b-3p could significantly reduce the thoracic aorta pathological damage in AD mice.
Conclusion: LncRNA H19 regulated smooth muscle cell function by sponging miR-193b-3p and it participated in the development of AD.
Aortic rupture/dissection (AD) is caused by an intima-media tear in the aorta under the impact of high velocity and pressure blood flow, forming a false or true lumen [1]. AD has a high mortality rate and a poor prognosis, and is a clinically urgent problem to be solved [2]. Thoracic AD (TAD) is a type of AD, according to pathological morphology [2]. Recent studies found that TAD is a comprehensive pathological change process caused by pathological changes involving multiple blood vessel constituents, such as human aortic smooth muscle cells (HASMCs) and extracellular matrix (ECM) [3,4].
Apart from the p53 suppression > H19 promotion > miR-193b-3p sponging axis, BA.5 subvariant specific mRNA booster agents also upregulate Ang II due to strong binding affinity to ACE2, so increased numbers of cases of CVD and AD are to be expected via 2 distinct pathways75:
In conclusion, the mutated S protein of Omicron BA.5 showed a 63-fold higher adhesivity to ACE2 receptors, with expected enhanced activity of Ang II and concomitant Ang1–7 deficiency. Finally, the higher adhesivity to ACE2 receptors by the S protein generated by new vaccines eventually built on the aminoacidic sequence of Omicron BA.5 might enhance the imbalance between Ang II overactivity and of Ang1–7 deficiency.
Basic and clinical research is urgently needed to investigate the clinical impact of dysregulated RAS axis on SARS-CoV-2 disease and vaccination. Moreover, the potentially detrimental impact of the interactions between S proteins (viral or vaccine-induced) and ACE2 and other angiotensinases (involved in the processing of Ang II to Ang1,7) remains to be determined [9]. New experimental and clinical data exploring the relationships between different mechanisms of Ang II cleavage and accumulation will be valuable in guiding the development of vaccines and other therapeutic strategies against SARS-CoV-2 pandemic [[14], [15]].
AD+Vector refers to the use of H19 plasmids created with pmirGLO luciferase vectorto quantitatively evaluate microRNA (miRNA) activity.
AD+shH19 refers to injection of AD mice with lentivirus carrying shH19, which noticeably up-regulated the miR-193b-3p level.
Greatest pathology was observed from the combination of the highest relative H19 expression and lowest miR-193b-3p levels.
HE staining results showed obvious vascular media degeneration in the AD group, whose muscle fiber assembly was disordered, and the middle membrane thickness in the thoracic aorta increased remarkably (Figure 4C).
Figure 4. Effect of lncRNA H19 silencing on AD animal models. (A,B) The expression levels of lncRNA H19 and miR-193b-3p in Sham, AD, AD + Vector, AD + shH19 groups were detected by qRT-PCR (n=6). GAPDH and U6 were used as internal reference genes. (C) The histopathological changes of each group were evaluated by HE staining (n=6). (D–F) qRT-PCR and Western blot were used to detect the levels of phenotypic differentiation markers α-SMA and SM22α and the expression of MMP-2 and MMP-9 in each group. GAPDH served as a control. Each experiment was repeated three times independently at least, and the results were the means ± SD. qRT-PCR: quantitative reverse transcription real time polymerase chain reaction. ***P<0.001 vs. Sham; ^^P<0.01, ^^^P<0.001 vs. AD + Vector. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7823186/
In summary, our research indicated that H19, as an miR-193b-3p sponge, regulates smooth muscle cell function and participates in AD vascular remodeling.Importantly, for the first time, we discovered that H19/miR-193b-3p is a new pathogenic pathway in AD.This provides a direction for further research on the role of H19 in AD and new theoretical support for the pathological mechanism of AD.
Vascular wall remodelling compensates for blood flow limitations caused by atherosclerotic plaque growth and other pathologies. Hypertension can also be a symptom of this. From a paper by Renna et al (2013)76:
Figure 1. Changes are predominantly in media-to-lumen ratio (M/L), changing the vessel wall width for increased muscle mass (Figure 1(A)) or in the reorganization of cellular and noncellular elements (Figure 1(B)). Another mechanism of remodeling mainly involves changes in the dimensions of the lumen (Figures 1(C) and 1(D)). In this case, the restructuring of the active components and cell signals does not result in significant changes in the dimensions of the vascular lumen. Another form of vascular remodeling is microcirculation rarefaction (Figures 1(E) and 1(F)). https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3736482/
Figure 2. Schematic representation for the adaptation of these changes in different pathologies, including structural changes to the intima layer that contribute to remodeling of the vascular wall. Thus, outward remodeling compensates for atherosclerotic plaque growth and delays the progression of blood flow limitation during stenosis, whereas during restenosis, intimal hyperplasia causes a narrowing of the lumen. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3736482/
I also discussed vascular rarefaction in this Substack:
Two case study reports were published in December 2022 by Ahmed Mohammed, MD and Bernardo Cortese, MD77, “Spontaneous” Coronary Artery Dissection After SARS-CoV-2 Messenger RNA Vaccination.
Elevated troponin is an indication that myocardial damage has occurred.
C-reactive protein (CRP) is made by the liver and is a marker for inflammation:
Case 1
A 58-year-old female patient was admitted to the emergency department on February 2022 with severe chest pain that had started 2 hours before presentation, occurring 2 days after SARS-CoV-2 messenger RNA (mRNA) vaccination (Comirnaty; Pfizer-BioNTech). A physical examination yielded unremarkable results. The patient had the following cardiovascular risk factors: arterial hypertension and dyslipidemia. Laboratory tests showed a troponin value of 3205 ng/mL (normal value [n.v.], <12 mg/mL) and yielded positive results for inflammatory markers such as C-reactive protein (CRP) (5.4 ng/mL; normal value, <0.5 ng/mL). Echocardiography showed preserved left ventricular ejection fraction with apical and septal hypokinesia, with no significant valvular defect. The patient was brought to the catheterization laboratory, where she was diagnosed with non–ST-elevation myocardial infarction. Proximal and middle left anterior descending artery showed a plaque determining 50% tandem stenosis, with impaired flow distally (Figure 1A), whereas the other coronary segments were normal. Because of the atypical angiographic pattern of such lesions, we decided to perform intravascular ultrasound imaging, which revealed a dissection of the left anterior descending artery into the ostioproximal segment, with an intramural hematoma (Figure 1B). Based on the SCAD European consensus document,3 because the vessel was well patent and the distal flow was maintained, the patient was treated conservatively with statins and a single antiplatelet agent. The patient was discharged 2 days later and was asymptomatic. After 3 months, no further events occurred.
Figure 1: (A) Coronary angiography showing stenosis in the proximal aspect of the left anterior descending artery, with suspicion of spontaneous coronary artery dissection (SCAD). (B) Intravascular ultrasound imaging of the proximal aspect of the left anterior descending artery showing a dissection flap, with an intramural hematoma. (C) Coronary angiography showing stenosis in the middistal aspect of the circumflex artery, with high suspicion of SCAD. (D) Optical coherence tomography analysis confirming the dual-lumen appearance of SCAD, with no clear sign of vessel rupture. LAD, left anterior descending artery; LCX, left circumflex artery. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9722217/
Leukocytosis means you have a high white blood cell count:
Case 2
A 48-year-old female patient was admitted to our catheterization laboratory in January 2022 for a 3-day-long chest pain that occurred 4 days after SARS-CoV-2 mRNA vaccination (Comirnaty). A laboratory analysis showed mild leukocytosis (11.500/mL) and positive CRP results (13.3 ng/mL; normal value, <0.5 ng/mL). Angiography showed a diffusely narrowed distal circumflex artery (Figure 1C), with no other significant coronary artery disease. Because of suspicion of SCAD, we performed an optical coherence tomographic analysis (Supplemental Video 1) and confirmed the diagnosis based on a clear 2-lumen vessel and the absence of atherosclerotic coronary artery disease (Figure 1D). Because of the patency of the vessel, conservative management was chosen. The patient was still asymptomatic 3 months later.
A computerized tomography (CT) coronary angiogram is an imaging test that looks at the arteries that supply blood to the heart. A CT coronary angiogram uses a powerful X-ray machine to produce images of the heart and its blood vessels. The test is used to diagnose a variety of heart conditions78.
It would be useful to have bioassay lncRNA analysis too, especially to confirm elevated H19 as many other factors can also elevate the risk of a SCAD occurring79:
Females. Though SCAD can occur in both men and women, it tends to affect women more than men.
Childbirth. Some women who have had SCAD have recently given birth. SCAD has been found to occur most often in the first few weeks after delivery, but it can also occur during pregnancy.
Fibromuscular dysplasia (FMD). This condition, which causes irregular growth of cells in artery walls, can weaken artery walls, leading to blockages, dissections or aneurysms. It can also cause high blood pressure, stroke and tears in other blood vessels. Women are more likely to have FMD than men.
Hormone use. Hormone therapy, such as from oral contraceptives or infertility treatments, has been associated with SCAD.
Other conditions affecting blood vessels. Diseases that cause inflammation of the blood vessels, such as lupus and polyarteritis nodosa, also have been associated with SCAD.
Inherited connective tissue diseases. Genetic diseases that cause problems with the body's connective tissues, such as vascular Ehlers-Danlos syndrome and Marfan syndrome, have been found to occur in people who have had SCAD.
Very high blood pressure. Severe high blood pressure can be associated with SCAD.
Illegal drug use. Using cocaine or other illegal drugs might increase the risk of SCAD.
Symptoms can include:
Chest pain
A rapid heartbeat or fluttery feeling in the chest
Pain in the arms, shoulders, back or jaw
Shortness of breath
Sweating
Unusual, extreme tiredness
Nausea
Dizziness
We shall now review some of the other lncRNAs that can potentially interact with Spike mRNA, as per earlier discussion:
LncRNA FENDRR
From a 2014 study by Xu et al and decreases expression of FENDRR is associated with poor prognosis in gastric cancer and FENDRR regulates gastric cancer cell metastasis by affecting fibronectin1 expression80. It appears to act as a tumour suppressor:
Results: FENDRR was downregulated in gastric cancer cell lines and cancerous tissues, as compared with normal gastric epithelial cells and adjacent noncancerous tissue samples. Low FENDRR expression was correlated with deeper tumor invasion (p < 0.001), higher tumor stage (p = 0.001), and lymphatic metastasis (p = 0.007). Univariate and multivariate analyses indicated that low FENDRR expression predicted poor prognosis. Histone deacetylation was involved in the downregulation of FENDRR in gastric cancer cells. FENDER overexpression suppressed invasion and migration by gastric cancer cells in vitro, by downregulating FN1 and MMP2/MMP9 expression.
LncRNA HOTAIR
A study by Rajagopal et al (2020) concluded that HOTAIR promotes tumour progression81:
Recent studies have identified that HOTAIR facilitates protein-protein interaction thereby affecting diverse pathways in cancer such as epigenetic reprogramming, protein stability and signal transduction. HOTAIR has been shown to promote tumor progression by regulating microRNA expression and function. Moreover, several HOTAIR gene variants have recently been identified and found to increase cancer susceptibility.
LncRNA MIAT
From Fasolo et al (2021) and upregulation of MIAT is positively associated with advanced atherosclerotic lesion formation and plaque destabilization82.
We profiled differences in RNA transcript expression in patients with advanced carotid artery atherosclerotic lesions from the Biobank of Karolinska Endarterectomies. The lncRNA MIAT was identified as the most upregulated noncoding RNA transcript in carotid plaques compared with nonatherosclerotic control arteries, which was confirmed by quantitative real-time polymerase chain reaction and in situ hybridization.
Conclusions: The lncRNA MIAT is a novel regulator of cellular processes in advanced atherosclerosis that controls proliferation, apoptosis, and phenotypic transition of SMCs, and the proinflammatory properties of macrophages, as well.
LncRNA APOA1-AS
Also pro-atherosclerosis, APOA1-AS was found to facilitate the proliferation and migration and to repress the apoptosis of arterial lining cells associated with coronary atherosclerosis. From Wang et al (2021)83:
Results demonstrated that APOA1-AS was notably up-regulated in ox-LDL treated VSMCs (ox-LDL-VSMCs). The deficiency of APOA1-AS hindered proliferation and migration and stimulated apoptosis in ox-LDL-VSMCs. Mechanistically, APOA1-AS recruited TATA-box binding protein associated factor 15 (TAF15) protein to stabilized SMAD family member 3 (SMAD3) mRNA and activate the TGF-β/SMAD3 signaling pathway. In conclusion, APOA1-AS contributed to proliferation and migration and repressed apoptosis of VSMCs through TAF15-mediated SMAD3 mRNA stabilization, indicating that APOA1-AS could be a promising target for CAS.
LncRNA LINC01505
A 2019 paper by Ye et al associates LINC01505 with neuroblastoma proliferation and apoptosis due its action as an oncogene84.
Neuroblastoma is a rare type of cancer that can affect children, usually under the age of 5, and it develops in specialised nerve cells left behind from a baby's development in the womb.
The 5-year survival rate for low-risk neuroblastoma is higher than 95%, for intermediate-risk neuroblastoma 90-95% and for high-risk neuroblastoma around 50%85:
Knockdown of linc01105 inhibited neuroblastoma cell proliferation, migration and invasion, and it induced apoptosis. In addition, linc01105 affected the expression of p53 and Bcl‑2 family proteins and activated the caspase signaling pathway. Further functional experiments revealed that linc01105 promoted the expression of the miR‑6769b‑5p target gene VEGFA by acting as a sponge of miR‑6769b‑5p. In conclusion, linc01105 may contribute to neuroblastoma tumorigenesis and development. The present findings indicated that the interplay between the p53/caspase pathway and the linc01105/miR‑6769b‑5p/VEGFA axis may have important roles in the development of neuroblastoma.
KEGG pathway analysis
Moving forward to 2022, and Enguita et al performed a study of a cohort of patients with different SARS-CoV-2 viral loads and analyzed the involvement of lncRNAs in supporting regulatory networks based on their interaction with RNA-binding proteins (RBPs)86.
What is of interest from their in silico analysis is that they also used experimental evidence to confirm their findings, by performing a transcriptomic analysis from nasopharyngeal swabs collected from patients testing for SARS-CoV-2 virus.
They found that a higher viral load correlated with a greater proportion of upregulated transcripts represented by lncRNAs, the main ones being NRIR, BISPR, MIR155HG, FMR1-IT1, USP30-AS1, and U62317.2.
From analysis of viral spike protein S mRNA for overexpressed lncRNAs they found moderately positive correlations with NRIR and BISPR. A low positive correlation was found with microRNA MIR155HG. We shall later explore pathologies associated with persistent overexpression of these.
A summary chart for all of these:
Figure 2. Functional prediction analysis by ncFANs 2.0 algorithm 39 of the upregulated lncRNAs observed in SARS-CoV-2 patients with high viral loads.A, GO-term enrichment analysis performed with ncFANs and filtered by removal of the redundant terms with REVIGO 41. The filtered GO-terms are classified according to their two-dimensional arbitrary semantic space and represented by symbols with dimensions proportional to the LogSize, showing the most relevant GO-terms for the context of viral infections. B, pathway enrichment analysis by ncFANs using the KEGG database. C, molecular signature analysis by ncFANs using the MSigDB database. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9131284/
Of note here are commonalities found via pathway enrichment analysis using the KEGG database. In decreasing order:
From molecular signature analysis by ncFANs using the MSigDB database:
Allograft rejection.
Epithelial mesenchymal transition94, which is related to healing, inflammation, cancer cell transformation, metastasis and cataract formation95.
Interferon gamma response. This relates to 2 upregulated lncRNAs in particular:
NRIR and BISPR. Again, its a log scale on the Y-axis:
Figure 3. Expression levels of selected lncRNAs quantified by next-generation sequencing in nasal swabs from the working group of patients, and whole blood, obtained from the COVIDOME project database74. LncRNA expression in nasal swabs distributed by groups of patients: A, NRIR; B, LINC02068; C, BISPR; D, USP30-AS1; E, MIR155HG and F, AL512306.2. LncRNA levels in whole blood in SARS-CoV-2 patients (Covid19) and non-infected controls (None): G, NRIR; H, BISPR; I, USP30-AS1 and J, AL512306.2. Statistical comparisons between sample groups were made by one-way ANOVA in the case of samples from nasal swabs and by the Student's t-test in the data from the COVIDOME project (****, p-value < 0.0001; ***, p-value < 0.001; **, p-value < 0.01; *, p-value < 0.05 and n.s, non-significant). https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9131284/
NRIR drives the interferon response in monocytes and is associated with systemic sclerosis (scleroderma)96 and Sjögren’s disease97.
BISPR is upregulated by interferon and acts as an antiviral98.
AL512306.2 is an m7G-related lncRNA which is associated with tumour immunity and poor prognosis100.
Analyzing the correlations of S-protein mRNA with the overexpressed lncRNAs, we found moderate positive correlation with NRIR (Figure (Figure4B) and BISPR (Figure (Figure4C) lncRNAs and low positive correlation with MIR155HG (Figure (Figure44D).
Figure 4. Spearman's correlation analysis of the top 90 upregulated lncRNAs and the viral gene transcripts in the cohort of analyzed samples.A, Hierarchical clustered Spearman's correlation matrix for the overexpressed lncRNAs and the detected SARS-CoV-2 transcripts across all the samples analyzed by the BioCPR software 75. The SARS-CoV-2 mRNA transcripts are highlighted within boxes; B, correlation analysis between NRIR lncRNA and S-protein transcript; C, correlation analysis between BISPR lncRNA and S-protein transcript; and D, correlation analysis between MIR155HG lncRNA and S-protein transcript. The correlation coefficients showed in panels b, c and d correspond to the Spearman analysis and are significant in all cases with p-values < 0.0001. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9131284/
The above analyses are derived from viral interactions. Antiviral interferon response is one of the most significant correlations to increased lncRNA expression of NRIR and BISPR, which is what you would want as a first line immune response, unless it is to excess.
In contrast to this, analysis from a 2022 by Seneff et al found that the interferon response in the mRNA transfected was somewhat suppressed101. We also saw this earlier with influenza vaccination.
Key highlights:
…In this paper, we present evidence that vaccination induces a profound impairment in type I interferon signaling, which has diverse adverse consequences to human health.
…Differential gene expression analysis of peripheral dendritic cells revealed a dramatic upregulation of both type I and type II interferons (IFNs) in COVID-19 patients, but not in vaccinees.One remarkable observation they made was that there was an expansion of circulating hematopoietic stem and progenitor cells (HSPCs) in COVID-19 patients, but this expansion was notably absent following vaccination. A striking expansion in circulating plasmablasts observed in COVID-19 patients was also not seen in the vaccinees. All of these observations are consistent with the idea that the anti-COVID-19 vaccines actively suppress type I IFN signaling,
…Type I IFNs play an essential role in fighting viral infections, and deficiencies in type I IFN signaling have been associated with poor outcomes from COVID-19 in multiple studies.
…Finally, Stertz and Hale (2021) note that, whether due to autoantibodies or perhaps loss-of-function polymorphisms associated with interferon system genes, deficiencies in interferon production are associated with as many as 15% of all life-threatening COVID-19 cases.
…In further exploration, the authors identified two microRNAs that were present in high concentrations in the exosomes: miR-148a and miR-590.They proposed a specific mechanism by which these two microRNAs would specifically disrupt type I interferon signaling, through suppression of two critical proteins that control the pathway: ubiquitin specific peptidase 33 (USP33) and IRF9.
…A study by de Gonzalo-Calvo et al. (2021) looked at the microRNA profile in the blood of COVID-19 patients and their quantitative variance based upon disease severity. Multiple miRNAs were found to be up- and down-regulated. Among these was miR-148a-3p, the guide strand precursor to miR-148a. However, miR-148a itself was not among the microRNAs catalogued as excessive or deficient in their study, nor was miR-590. It appears from these findings that miR148a and miR-590 and their inflammatory effects are unique to vaccination-induced SARS-CoV-2 spike glycoprotein production.
…A central point of our analysis below is the important distinction between the impact of vaccination versus natural infection on type I IFN. While vaccination actively suppresses its production, natural infection promotes type I IFN production very early in the disease cycle. Those with preexisting conditions often exhibit impaired type I IFN signaling, which leads to more severe, critical, and even fatal COVID-19. If the impairment induced by the vaccine is maintained as antibody levels wane over time, this could lead to a situation where the vaccine causes a more severe disease expression than would have been the case in the absence of the vaccine.
Another expected consequence of suppressing type I IFN would be reactivation of preexisting, chronic viral infections, as described in Section 9.
Impaired type 1 interferon signalling means that latent viruses like herpes (Varicella zoster) can be reactivated, leading to shingles. NEAT1 suppression would enhance this effect, as discussed previously.
And if it is an oncogenic virus such as HPV this can also lead to increased risk of carcinogenesis and a strong association has been found between reactivated viruses and onset of aggressive breast cancer through competition for a BRCA1 precursor called GABP. See Polansky & Schwab (2019)102.
Unique to the transfected, miR590 was being upregulated. From Zhou et al (2018)103:
MicroRNA (miR)-590-5p has been identified as an important regulator of some signaling pathways such as cell proliferation and tumorigenesis. However, little is known about its role during viral infection. Here, we report that miR-590-5p was significantly induced by various viruses and effectively potentiated virus replication in different viral infection systems. Furthermore, miR-590-5p substantially attenuated the virus-induced expression of type I and type III interferons (IFNs) and inflammatory cytokines, resulting in impaired downstream antiviral signaling. Interleukin-6 receptor (IL6R) was identified as a target of miR-590-5p.
As we saw earlier, suppression of NEAT1 and upregulation of MALAT1 could also compound the immunosuppression and reactivation which is not coincidental as NEAT1 acts as a sponge to reduce levels of miR-148a as it has a binding site. Macrophage skewing to inflammatory M1 from M2 also correlates with NEAT1 inhibition and miR-148a upregulation, as does upregulation of MALAT1.
Additionally, lncRNA NEAT1 increased in CNV formation and M2 macrophage polarization. LncRNA NEAT1 sponging miRNA-148a-3p targeting PTEN can modulate M2 macrophage polarization in mice CNV models as well as in bone marrow-derived macrophages cultured in vitro. Inhibition of lncRNA NEAT1 can suppress M2 macrophage both in vitro and vivo.
…Moreover, miR-148a-3p also enhanced M1 macrophage polarization and pro-inflammatory responses through PTEN/AKT-mediated upregulation of NF-κB signaling. In summary, our data establish a novel molecular mechanism by which Notch signaling promotes monocyte differentiation and M1 macrophage activation through miR-148a-3p.
NEAT1 and immunosuppression due to apoptosis of hematopoietic stem cells
The Seneff paper made reference to “..an expansion of circulating hematopoietic stem and progenitor cells (HSPCs) in COVID-19 patients, but this expansion was notably absent following vaccination”.
Circulating HSPC’s are important precursor cells of the immune system and support blood and tissue regeneration.
Only about 1 in 10,000 cells in myeloid tissue are HSC’s, so a loss of only a few of these can have disproportionally large downstream effects on the immune system and erythrocyte generation, with long recovery periods if the damage is severe enough.
Your HSC’s do not have an unlimited lifespan, even if that period extends out several years. From Sieburg et al (2013)106:
The life-long supply of blood cells depends on the long-term function of hematopoietic stem cells (HSCs). HSCs are functionally defined by their multi-potency and self-renewal capacity. Because of their self-renewal capacity, HSCs were thought to have indefinite lifespans. However, there is increasing evidence that genetically identical HSCs differ in lifespan and that the lifespan of a HSC is predetermined and HSC-intrinsic. Lifespan is here defined as the time a HSC gives rise to all mature blood cells.
…Clonal hematopoiesis begins with a single HSC and ends with its loss after months to years [4]–[8]. During this period of time - the lifespan -, the genetic/epigenetic program of the original HSC is replicated to many daughter HSCs. In HSC self-renewal, discrepancies between replicates should be vanishingly small. Yet, clones extinguish after a limited time, suggesting that self-renewal may not be perfectly reliable in the long-run.
lncRNAs don’t just affect NOTCH signalling, monocyte differentiation and M1 macrophage activation. There are also implications for carcinogenesis and NEAT1 appears to be at the heart of this with respect to HSC pathology.
NOTCH signalling pathology alone is a subject worthy of its own dedicated future Substack.
Notch signaling can elicit opposite effects in regulating tumorigenicity (tumor-promoting versus tumor-suppressing function) as well as controlling immune cell responses. In various cancer types, Notch signaling elicits a "cancer stem cell (CSC)" phenotype that results in decreased proliferation, but resistance to various therapies, hence potentially contributing to cell dormancy and relapse. CSCs can reshape their niche by releasing paracrine factors and inflammatory cytokines, and the niche in return can support their quiescence and resistance to therapies as well as the immune response. Moreover, Notch signaling is one of the key regulators of hematopoiesis, immune cell differentiation, and inflammation and is implicated in various autoimmune diseases, carcinogenesis (leukemia), and tumor-induced immunosuppression. Notch can control the fate of various T cell types, including Th1, Th2, and the regulatory T cells (Tregs), and myeloid cells including macrophages, dendritic cells, and myeloid-derived suppressor cells (MDSCs). Both MDSCs and Tregs play an important role in supporting tumor cells (and CSCs) and in evading the immune response.
TGFb and CD34+ are implicated in this too, with far reaching consequences as a 2022 study by Estep et al found that HSCs in cord blood was also being depleted, with consequences for the neonate potentially including immunosuppression or failure to carry full term108.
Two independent hematopoiesis assays, a colony forming unit assay and a mouse humanization assay, revealed skewed hematopoiesis of HSPCs obtained from donors delivered from mothers with SARS-CoV-2 infection history. These results indicate that SARS-CoV-2 infection and COVID-19 vaccination impair the functionalities and survivability of HSPCs in the UCB, which would make unprecedented concerns on the future of HSPC-based therapies.
…More importantly, UCB is highly enriched with multipotent hematopoietic stem progenitor cells (HSPCs) as identified by the surface expression of CD34 molecules, which are essential for the maintenance of the bone marrow and blood systems
…These data indicate that both previous SARS-CoV-2 infection and/or vaccination impair CD34+ HSPCs quantitatively and qualitatively by stress-induced hematopoiesis, which is a great concern in the collection as well as the utilization of UCB as a source of CD34+ HSPCs used in/for future therapies, treatments, and research.
…The decrease and the stress-induced differentiation of CD34+ HSPCs observed attributes to decreased circulating peripheral lymphocytes in fetuses, resulting in lower MNC numbers in the UCB. At present, it is unknown how these damages on CD34+ cells are impactful to the neonate immune system for example, for fighting other pathogenic viruses, bacterial infections, or cancerous cells, as well as for maintaining homeostatic hematopoiesis, proper humoral and cellular immunities.
The authors do not refer to Notch signalling pathways. Instead they suspect depletion of CD34+ cells is being caused by continuous IFN-γ stimulation, with prior infection being a key factor.
Notch signalling pathways are highly complex and considered in detail in this 2007 paper by Chadwick et al.
They performed an in vitro study into how Notch signalling induces apoptosis in CD34+ HSPCs109:
The results of this study show that activation of the Notch signaling pathway has an inhibitory effect on the proliferation and survival of human hematopoietic CD34+ cells populations. These findings have important implications for strategies aimed at promoting self-renewal of human HSCs.
…In this study, we have analyzed the functional effect of Notch signaling on human hematopoietic progenitor cells (including the CD34+Lin−Thy+ HSC compartment), by ectopic expression of constitutively active Notch 1. We show that ectopic expression of N1ICD leads to cell cycle arrest and apoptosis in CD34+ cell populations in a mechanism that may be mediated by the upregulation of p21 and BCL2L1.
Transforming growth factor beta (TGF-β) induced apoptosis of HSPCs
There is some overlap in some of the pathways being discussed, eg interferon signalling but TGF-β itself can also induce bone marrow failure syndromes, which is characterised by ineffective hematopoiesis due to impaired fitness of hematopoietic stem cells. Javier et al (2021) conducted various assay analyses and used a mouse model to investigate further110:
In this study, we used a transgenic conditional mouse model over-expressing constitutively active TGFb1 (aTGFb1)19 to further investigate the role of TGFb1 in BMF/MDS. We show that a physiological increase in aTGFb1 production in the bone marrow only produces mild neutropenia and anemia in mice during aging. However, the combination of increased TGFb signaling plus polyinosinic:polycytidilic acid (pIC)-driven acute inflammatory stress drives chronic BMF with phenotypes similar to those of the human disorders associated with ineffective hematopoiesis, including BMF syndromes and MDS. Mechanistically, TGFb prevents the termination of an acute pIC response causing permanent alteration in mitochondrial functions and increased caspase-1 activity. Our findings therefore suggest that BMF syndromes can be initiated solely by multiple inflammatory hits in the context of increased TGFb signaling, and that disease outcome is dependent on the inflammatory context. Increased TGFb signaling plus pIC thus represents a novel non-genetic-driven mouse model of human BMF-like diseases.
The link with exposure to spike protein from either infection or experimental gene therapy agents is that depletion of ACE2, as discussed earlier, can induce elevated TGFb1 levels. From a 2012 investigation into TGF-β/Smad-mediated renal fibrosis by Liu et al111:
In conclusion, enhanced Ang II-mediated TGF-β/Smad and NF-κB signaling may be the mechanisms by which loss of Ace2 enhances renal fibrosis and inflammation. Smad7 ubiquitin degradation mediated by Smurf2 may be a central mechanism by which Ace2−/y mice promote TGF-β/Smad2/3-mediated renal fibrosis and NF-κB-driven renal inflammation in a mouse model of UUO nephropathy.
…It has been shown that Ang II is capable of activating the Smads to mediate fibrosis via both AT1-MAP kinase-crosstalk and TGF-β1-dependent pathways.15, 16, 17 We thus investigated whether enhanced renal fibrosis in the UUO kidney of Ace2−/y mice is associated with an increase in Ang II-mediated TGF-β/Smad pathway. As shown in Figure 4a and b, significant higher levels of intrarenal Ang II and Ang 1–7 were detected in Ace2+/y mice after UUO. Deletion of Ace2 resulted in doubling the intrarenal Ang II in the UUO kidney, while levels of intrarenal Ang 1–7 remained low
…Further study revealed that enhanced Ang II signaling in the UUO kidney of Ace2−/y mice was accompanied by a marked increase in renal TGF-β1 expression (Figure 5a, c and d) and higher levels of Smad2/3 phosphorylation and phospho-Smad2/3 nuclear translocation when compared with Ace2+/y mice (Figure 5b and e–g).
In summary:
1/ Interferon mediated:
Spike mRNA > IFN-γ > CIITA mediated HLA-class II gene expression > various effects on CD34+ HSPCs: apotosis/disrupted quiescence/excessive terminal differentiation/negative impacts on multilineage engraftment & renewability
2/ Via NEAT1 suppression & NOTCH pathways:
Spike mRNA > NEAT1 suppression > less sponging of miRNA-148a-3p > NOTCH signalling > CD34+ HSPC apoptosis
3/ Via NEAT1 upregulation & inflammasomes:
Severe COVID-19 disease > NEAT1 upregulation > activation of NLRP3, NLRC4 and AIM2 inflammasomes > pyroptosis of HSPCs
4/ Via TGF-β signalling:
Spike mRNA > ACE2 depletion > Ang II upregulation > NF-kB/TGF-β/Smad phosphorylation > pIC driven inflammation > mitochondrial damage/caspase-1 > chronic bone marrow failure
Bone marrow HSPC recovery periods
These studies aren’t directly comparable as different stem cells may be affected in different ways, but they are useful for guidance as to how drawn out recovery can be.
In a paper from 1989 by Casamassima et al they used magnetic resonance imaging (MRI) to assess hematopoietic bone marrow recovery periods after radiotherapy. They concluded that partial recovery took 2 - 9 years and and full recovery could take as long as 10-23 years112.
At lower doses 1 year may elapse before significant hematopoietic tissue injury is apparent.
And after stem cell or bone marrow transplant it can take 6 to 12 months for blood counts to return to normal, bearing in mind that this is with healthy donor cells113.
Further NEAT1 & MALAT1 related pathologies
NEAT1 also binds (ie sponges) inflammatory, interferon suppressing miR-590.
Further experiments revealed that NEAT1 could sponge miR-590-3p. Knockdown of miR-590-3p reversed the inhibitory effect of NEAT1 knockdown on LPS-induced H9c2 cell damage. Additionally, the NEAT1/miR-590-3p axis could regulate the activity of the NF-κB signaling pathway. To conclude, lncRNA NEAT1 accelerated apoptosis and inflammation in LPS-stimulated H9c2 cells via sponging miR-590-3p. These findings may provide a new strategy for the treatment of sepsis.
The evidence points to suppression of NEAT1 by mRNA transfection in contrast to the effects of SARS-CoV-2 infection on the un-transfected or un-boosted, thus leading to elevated levels of interferon suppressing miR-148a and miR-590 and a skewed inflammatory M1 type macrophage response, which would be more pathogenic to tissues, but with suppressed antiviral properties.
Less paraspeckles also promotes viral reactivation.
A literature search shows that MALAT1 also sponges various mRNAs, but not miR-148a. It does, however inhibit miR-590-3p according to an investigation of its effects on osteosarcoma ("OS", bone cancer)115. This would be in opposition to the effects of NEAT1, if occurring in the same tissues at the same time and to the same degree. Quantification of the in vivo effects of transfection on both of these miRNAs and interferon therefore requires further studies.
The results indicated that MALAT1 was highly expressed in OS tissues and cell lines. MALAT1 knockdown promoted apoptosis and suppressed proliferation, migration, invasion and epithelial- mesenchymal transition (EMT) of OS cells. Overexpression of miR-590-3p increased cell apoptosis and hampered cell proliferation, migration, invasion and EMT in OS cells. In addition, MALAT1 knockdown upregulated the expression of miR-590-3p in OS cells. In conclusion, MALAT1 was demonstrated to suppress cell apoptosis and induce cell proliferation, migration, invasion and EMT by inhibiting miR-590-3p in OS, which indicated that MALAT1 has potential value in the diagnosis and treatment of OS.
A Bill & Melinda Gates Foundation funded study from November 2022 by Chattopadhyay et al admits to its shortcomings but did indeed find that in the samples analysed several lncRNAs were being downregulated in the vaccine breakthrough group (VBT), including NEAT1 & MALAT1116. This led to downregulation of innate immune and inflammatory response in the vaccination breakthrough infection group, coupled with a higher viral load and therefore a longer time to clearance.
There are also questions to be asked about suppressed immune responses to other infections too, as well as cancer, as discussed previously.
The lncRNA-mediated suppression of innate immune/inflammatory responses and activation of adaptive immune response might explain the SARS-CoV-2 breakthrough infections with milder symptoms in the VBT.
…prior research by our lab and others has emphasized the function of lncRNAs in the SARS-CoV-2 infection and how it influences differential disease severities (8–10). Therefore, we conducted this first-of-its-kind study to understand the role of lncRNAs in the vaccination breakthrough infection, which might explain why despite the vaccination, the individuals were infected with the SARS-CoV-2.
…Out of the 727 DE-lncRNA transcripts, 32 lncRNA transcripts, corresponding to 15 lncRNA genes have known functions associated with the immune/inflammatory response. Further, out of the 12 lncRNA genes, 7 lncRNAs are reported to modulate immune/inflammatory responses in infectious diseases. Out of the 7 lncRNAs, LUCAT1, MALAT1, NEAT1 and GAS5 were downregulated in the VBT.LUCAT1 is a negative regulator of interferon response, and downregulation of LUCAT1 suggests an activated interferon response.
…The patients were stratified into two groups based on their vaccination status; Vaccination Breakthrough (n=28) with ChAdOx1 nCoV-19 vaccination prior to infection and Unvaccinated (n=29) including patients without any prior vaccination.
…The clinical data of the vaccination breakthrough patients highlight that despite having higher viral load, they have milder symptoms compared to the unvaccinated COVID-19 patients.
…While we highlight the above findings, it is important to highlight the possible ways of future strengthening of the findings. The study is based on the nasopharyngeal RNA collected from the COVID-19 patients at the day of hospital admission/home quarantine. While this helps understanding the initial host response at the site of entry, availability of blood samples, and samples post-hospitalization could have increased our understanding of the adaptive immune response. Besides, the neutralizing antibodies could not be measured due to unavailability of the blood samples.
This meant that although at the time the patients appeared or felt less ill in comparison to the unvaccinated group underlying viral induced damage could, repeatedly, cause further pathologies in the longer term. These would typically be seen as old age diseases: cardiovascular disease, cancers, T-cell anergy, HSC depletion, neurodegeneration, autoimmune disorders, mitochondrial & metabolic dysfunction, kynurenine pathway promotion, endocrine effects and so on.
They also found that LINE1 was significantly higher in their interacting lncRNAs, which also elevates the risk of tumorigenesis and is the hallmark of many cancers117.
The LINE1 was also significantly higher in the interacting lncRNAs (p value < 0.0001), however, the majority of the LINE1 was present in the antisense strand. Alus are known to modulate immune response by homologous interaction with the immune responsive genes, and to activate interferon response (48, 49). While some studies highlight the accumulation of LINE1 upon viral infection, LINE1 is reported to activate the antiviral response in specific instances (50, 51). Besides, the antisense LINE1 is reported to regulate the expression of surrounding genes (52). The LTRs help in viral replication, and it is also reported to activate host immune response (53, 54). Since the interacting lncRNAs are expressed in the VBT group, the higher abundance of Alu, LINE1, and LTR elements within the interacting lncRNAs suggest a possible regulation of antiviral and immune response by the interacting lncRNAs in the vaccination breakthrough individuals.
And in 2017 Mello et al used knockout mice in a study of the tumour suppressing properties of NEAT1118. Downregulation of innate immune and inflammatory responses, especially due to downregulation of NEAT1 in the VBT group, has further implications in terms of elevated cancer risk or risk of recurrence:
Using fibroblasts derived from Neat1−/− mice, we examined the functional role of Neat1 in the p53 pathway. We found that Neat1 is dispensable for cell cycle arrest and apoptosis in response to genotoxic stress. In sharp contrast, Neat1 plays a crucial role in suppressing transformation in response to oncogenic signals. Neat1 deficiency enhances transformation in oncogene-expressing fibroblasts and promotes the development of premalignant pancreatic intraepithelial neoplasias (PanINs) and cystic lesions in KrasG12D-expressing mice. Neat1 loss provokes global changes in gene expression, suggesting a mechanism by which its deficiency promotes neoplasia. Collectively, these findings identify Neat1 as a p53-regulated large intergenic ncRNA (lincRNA) with a key role in suppressing transformation and cancer initiation, providing fundamental new insight into p53-mediated tumor suppression.
What regulates the master regulators?
Two mechanisms are in play:
Transcription factors.
Binding sites for Sox2, Oct4 and Nanog have been found on several lncRNA promotors. Sox2 (SRY-box transcription factor 2), is a gene that provides instructions for making a protein that plays a critical role in the formation of many different tissues and organs during embryonic development.
SOX2 expressed protein is especially important for the development of the eyes119. The Oct4 gene helps to maintain and regaining stem cell pluripotency120.
And the Nanog gene performs much the same: it is involved in embryonic stem (ES) cell proliferation, renewal, pluripotency as well the autorepression of its own expression in differentiating cells121.
Most striking is that lincRNA-p21 is one of many LncRNAs whose promoter regions contain binding sites for p53. P53 mediated expression of p21 from lincRNA-p21 is necessary to induce apoptosis in response to DNA damage, thus acting as a tumor suppressor itself.
…multiple evidence has reported that, lincRNA-p21 involves in the development of different tumors. Accordingly, the lincRNA-p21 overexpression can suppress tumor invasion by Notch pathway [6]. Also, lincRNA-p21 suppresses the progression of prostate cancer by apoptotic induction and controlling gene expressions downstream of p53 [7]. In fact, it plays a significant role as a moderator in different processes such as DNA damage response, apoptosis, and in cell proliferation.
These include histone methylation and DNA methylation.
Examples of epigenetic changes induced by methylation include due to smoking cigarettes122.
Histone methylation is the modification of certain amino acids in a histone protein by the addition of one, two, or three methyl groups. In the cell nucleus, DNA is wound around histones. Methylation and demethylation of histones turns the genes in DNA "off" and "on,” respectively, either by loosening their tails, thereby allowing transcription factors and other proteins to access the DNA, or by encompassing their tails around the DNA, thereby restricting access to the DNA.
From a featured review on the arraystar.com site, the excellently titled Wake Up and Smell the Chromatin Regulation of LncRNA Gene Expression123:
Regulation of LncRNA gene expression
Recently, increased attention has been focused on the transcriptional regulation of the LncRNA genes themselves. LncRNAs are transcribed by RNA polymerase II, and most of them have the hallmarks typical of pol II-transcribed gene products: 7-methylguanosine capping and polyadenylation. Many LncRNAs are expressed in tissue- and/or developmental specific patterns5. Therefore, the transcription of the LncRNA genes must be tightly controlled. As is typical for RNA polymerase II-transcribed protein-coding genes, LncRNA gene expression can be regulated by several different mechanisms (Figure 2).
Figure 2. LncRNA expression can be regulated by transcription and epigenetic factors. Blue coiled line represents an lncRNA promoter. Red coiled line represents an RNA polymerase II-transcribed, polyadenylated lncRNA (although not all lncRNA transcripts are polyadenylated). Bent arrow indicates direction of transcription. lncRNA transcription has been shown to be positively regulated by several well-characterized transcription factors (such as Oct4, Nanog, and p53). Epigenetic factors can have opposing effects on lncRNA transcription. Cytosine methylation (such as at CpG islands) can repress lncRNA expression, whereas methylation of lysine residues on histones (such as trimethylation of lysine 4 on histone H3) can activate lncRNA transcription.
Control by transcription factors
Recently, LncRNA genes have been shown to be targets of a number of well-characterized transcription factors. Several LncRNA promoters were found to contain putative binding sites for Sox2, Oct4 and Nanog6-8. Treatment of mouse embryonic stem cells (mESCs) with the differentiation factor retinoic acid causes the downregulation of Oct4 and Nanog RNA levels, as well as a concomitant decrease and increase of the LncRNAs AK028326 and AK141205, respectively.Subsequent RNA interference of Oct4 and Nanog led to similar effects on the expression levels of these LncRNAs.Further, Oct4 was found to be necessary for the transcription of no fewer than three LncRNAs in ESCs and induced pluripotent stem cells (iPSCs)8. Similarly, it was recently shown that lincRNA-p21 is one of many LncRNAs whose promoter regions contain canonical binding sites for the tumor suppressor protein p537,9. lincRNA-p21's transcription is directly induced by p53 in response to DNA damage, and loss of this induction leads to a loss of repression of many downstream p53 target genes. lincRNA-p21 was further shown to be necessary for p53's role in DNA damage-dependent apoptosis9. Therefore, the regulation of LncRNA expression is a critical factor associated with the initiation and development of many human cancers.
Control by epigenetic mechanisms
Several groups have further demonstrated that LncRNA expression levels are regulated by epigenetic mechanisms, including histone modificationas well as DNA methylation. In a large screen7, 1,675 mouse lincRNA genes in ESCs, embryonic fibroblasts (MEFs), neural progenitor cells (NPCs), and lung fibroblasts (MLFs) were identified on the basis of harboring a "chromatin signature": Histone H3 Lysine 4 trimethylation in promoter regions, and Histone H3 Lysine 36 trimethylation in the body of the gene. These so-called "K4-K36 domains" are also found in protein-coding genes and are therefore excellent predictors of the occurrence of transcribed units. In another study10, it was shown that the promoters of a subset of LncRNAs expressed in embryonic stem cells (ES cells) occur in regions for Histone H3 Lysine 4 and Histone H3 Lysine 27 trimethylation (H3K4me3 and H3K27me3, respectively). However, Many of these " bivalent marks" were lost in more differentiated NPCs and fibroblasts.
DNA methylation at CpG dinucleotides also plays a significant role in regulating the expression of LncRNA genes. In a lncRNA profiling study, microarray analysis was used to determine which LncRNAs were upregulated in the human colorectal cancer cell line HCT116 following treatment of these cells with the DNA demethylating agent 5'-aza-2'-deozycytidine. It was revealed that the expression of several lncRNAs are significantly upregulated due to the demethylation at the CpG islands in their promoters following exposure to the drug, indicating that the expression of these lncRNAs are controlled by their promoter DNA methylation level., In addition, two groups12,13 found silencing of expression of LncRNA Glt2 (Meg3) in both mouse induced pluripotent stem cells (iPSCs), as well as human hepatocellular cancer (HCC). In the HCC model, MEG3 expression is indirectly controlled by a micro RNA, mir-29a, which regulates the expression of methyltransferases necessary for DNA methylation, and inhibition of DNA methyltransferase activity in HCC cells causes de-repression of MEG3 expression. Together, these results suggest that the mechanism of regulation of the expression of a LncRNA in two disparate systems is conserved.
Summary
In summary, recent experimental results from several groups have revealed the coordinated regulation of expression of the long non-coding RNAs in a plethora of cell types. These studies have uncovered a major role of the epigenetic regulation of the LncRNA genes, opening up new possible therapeutic targets of the pathways involved with this novel class of non-protein coding RNAs, in many diseases including cancer.
RNA prions and lncRNA induced prionopathies
Hat tip to Maples for posting this link124. The paper was published by Mushegian and Elena in 2020 and poses the opinion/hypothesis that the definition of a prion (proteinaceous infectious agent) should include RNAs with these properties, not just proteins125:
Can there be RNA prions? We propose to take another step in expanding the definition of a prion and admit the possibility of the prion-like behavior in another class of biopolymers, namely, RNA. Even though RNA for a long time has been seen primarily as a carrier of genetic information or facilitator of protein synthesis, it is now clear that RNA molecules may perform catalytic and regulatory functions that do not involve encoding proteins but instead rely on the enzymatic activity, ligand-binding ability, or capacity for dynamic structural rearrangements of RNA itself.
…Most criteria for a protein-based prion can be generalized for RNA (Table 1, “RNA prion definition in this work”): an RNA prion is encoded by the cellular gene but inherited extrachromosomally; it has a phenotype that is due to the function of the RNA itself, not of its encoded protein, if such protein exists; and the phenotype mediated by an RNA prion is inducible and transmissible. RNA prions have two forms, a benign and a phenotype-causing one; analogously to the case of the protein prion [β], conversion between the benign and prion form of RNA may involve RNA processing. The benign form may undergo a rare conversion to the prion form, perhaps stimulated by stress or other external factors. When the prion form is already present, the rate of conversion of benign copies to prion form increases.
Several of the above properties have been observed in some naturally occurring or computationally designed RNA molecules.
…In addition, the analysis presented above immediately suggests several new modalities with interesting properties, which may be constructed and tested by synthetic biologists.
An interesting paper from 2018 by Russian researchers Mustafin and Khusnutdinova takes this further and hypothesises that the expression of specific lncRNAs may catalyse the transition of a prion protein (Prp C, C = common) to a prion confirmation (PrP Sc, S = scrapie).
A positive feedback mechanism is also proposed whereby miRNAs may, eventually, further upregulate lncRNA expression of the catalytic lncRNA in an exponential chain reaction126:
It is apparent that the probability of such events is extremely low, but if it occurs, the pathological process will be exponential, which is confirmed by both prion-disease epidemiology and their development pattern (long incubation period followed by fast development).
TSEs: Transmissible spongiform encephalopathies.
In the brain tissues of patients with prion diseases, as well as in exosomes containing an abnormal PrPSc isoform, changes in the levels of microRNA have been observed. A possible cause is the interaction of the spatial domains of PrPSc with the sequences of the non-coding RNA genes, which causes a change in their expression. MicroRNAs, in turn, affect the synthesis of long non-coding RNAs. We hypothesize that long noncoding RNAs and possibly microRNAs can interact with PrPC catalyzing its transformation into PrPSc. As a result, the number of PrPSc increases exponentially. In the brain of animals and humans, transposon activity has been observed, which has a regulatory effect on the differentiation of neuronal stem cells. Transposons form the basis of domain structures of long non-coding RNAs. In addition, they are important sources of microRNA. Since prion diseases can arise as sporadic and he-reditary cases, and hereditary predisposition is important for the development of pathology, we hypothesize the role of individual features of activation of transposons in the pathogenesis of prion diseases.
Amyloid fibrils are a structural state taken by polypeptides in relatively high concentrations in case they are not able to form more complex structures such as folds for a globular ferment structure or participate in a functional supramolecular complex (Tycko, 2014).
Formation of amyloids presents certain interest for medical studies, because the tissues contaminated by such amyloid diseases as T2D, neurodegenerative diseases (AD ,PD and TSE), accumulate amyloid fibrils (Tycko, 2014). For instance, TSEs are characterized by the infectious properties of the amyloid proteins causing the disease, while the encephalopathies themselves are described as a group of neurodegenerative diseases affecting the nervous system of humans and animals (Saba et al., 2012). In humans TSE causes such conditions as Creutzfeldt-Jacob disease (CJD), Gerstmann–Sträussler–Scheinker syndrome (GSS).
Multiple studies have demonstrated that the N-end domain can interact with a wide spectrum of ligands such as β-layer rich conformers (in particular PrPSc in TSE, and the Aβ42 peptide in AD); metal ions (Cu2+ and Zn2+); lipids; glycosaminoglycans (give different, sometimes opposite activity properties to the protein including protection from oxidative stress and PrPSc toxicity mediation); and nucleic acids (Eigenbrod et al., 2017). It also can be assumed that the changed conformation of PrPSc results in N-end domain disorder modulating its physiological function and enabling it to affect the genes of noncoding RNAs (ncRNA) by interaction with regulatory sequences or their gene regulators activating them in the way the peptides formed while ncRNA translation do. In rare cases, the activated ncRNAs can be a cofactor for the conformational transformation of PrPC into PrPS , and if it happens, it causes a chain reaction with the exponential growth of PrPSc, which is typical for prion diseases. However, the probability of such activation is negligibly small due to the long incubation period typical for prion diseases.
Apart from PrPSc, there are other elements that can either affect TSE development or determine the clinical phenotype of the disease and susceptibility to a prion infection, e.g. it has been found that PrPC- coding allelic PRNP can participate in TSE pathogenesis. It has also been found that the polymorphism of the ZBTB38 gene RASA2 (RAS p21 protein activator 2) is associated with susceptibility to CJD in the UK, while in China an expressed association has been confirmed between SNP rs57095329 in miR-146a and susceptibility to FFI (Gao et al., 2018). Studying TSE microRNAs as infection diagnostic markers in animals consumed as food has been considered as a perspective direction of research, e.g. in recent studies they used quantitative PCR with inverse transcription to analyze the levels of candidate microRNAs in the blood plasma of scrapie-infected sheep. In this study, a significant association between the disease and the high levels of miR-342-3p и miR-21-5p has been indicated (Sanz Rubio et al., 2017).
It has also been demonstrated that in TSE hippocampal neurons suffer microRNA deregulation (Burak et al., 2018), and this is a structure with the highest TE activity, which is assumed to be the basis of neural heterogeneity that plays the pivotal role in brain formation. This may lead to a conclusion that the peculiarities of the certain RNAs related to TEs through their brain regulation function can play their role in TSE development. Such a peculiarity can be accessibility of the certain genome sections containing TE sequences and microRNA genes that allow the PrPSc domains to interact with them. The result can be enhancement of the specific RNAs, a cascade of interacted processes develop-ing in geometric progression that affects the expression of the certain lncRNAs, which, in their turn, may interact with the PrPC isoform catalyzing its transition into PrPSc. It is not unlikely that microRNAs themselves can interact with prions as cofactors causing their conformation transformation. It is apparent that the probability of such events is extremely low, but if it occurs, the pathological process will be exponential, which is confirmed by both prion-disease epidemiology and their development pattern (long incubation period followed by fast development). Since every animal species has its own TE composition and positioning in the genome with its unique effect on the epigenetic properties of its functioning (Мустафин, Хуснутдинова, 2017), the described mechanism of TSE pathogenesis corresponds to the species-specific prion infection pattern.
To describe a possible mechanism of how a lncRNA can catalyze the conformation transformation of a common protein into a prion one should consider the similar mechanisms of prion-like domains (PLDs). PLDs enable for the ‘functional reaction’ of a protein that forms complexes of higher order and microscopic RNP granules. It is considered that protein and RNP concentration in confined space make gene regulation more effective. The biophysical properties of PLD-mediated interactions may explain the liquid-like properties of the RNP granules, as the PLDs expressed in vitro form hydrogels, so a PLD is the sequences with a low degree of complexity found the RNA-binding proteins associated with neurodegenerative amyotrophic lateral sclerosis (ALS).
The importance of NEAT1, paraspeckles and prion-like domains:
Many PLDs in proteins are binded with paraspeckles, sub-nuclear bodies formed around lncRNAs. Protein RBM14 binds the key paraspeckle subcomplexes via their mediated PLDs. It takes PLD proteins RBM14 и FUS to form paraspeckles in a cell, where their endogenous copies have been knocked down. They form hydrogel with amyloid-like properties. PLDs launch the liquid-phase transformations, which underlines the importance of this body as a model for understanding of neurodegeneration (Hennig et al., 2015)
Paraspeckle occurrence is initiated by NEAT1 transcription followed by enrichment with the different proteins that coordinate paraspeckle accumulation. The genes encoding paraspeckle protein are FUS, TDP-43, SS18L1, HNRNPA1, TAF15, EWSR1. Paraspeckles are sensitive to the stress caused by the structures induced by viral infections; by proteasomes and differentiation. They affect gene expression via keeping RNA in a nucleus through inverted repeats and sequestration of certain transcription factors (Hennig et al., 2015). Experiments in vitro have also demonstrated there is a synergism between RNA effects and PrPSc amplification. In this study, these were ultrasound-formed small RNA fragments that most easily converted into prions (Gonzalez-Montalban et al., 2011), which allows one to assume that microRNAs may serve as catalysts for PrPC transformation into PrPSc. Another study has also demonstrated that RNAs act as cofactors for formation of high-level de novo infectious prions facilitating PrP refolding into a pathogenic isoform (Timmes et al., 2013).
This has been demonstrated experimentally:
Searching for the factor facilitating PrPSc infectivity was carried out in (Simoneau et al., 2015). The experiments showed that initially nonpathogenic recPrP similar to PrPC could initiate a prion disease in hamsters via its prion-like conformation (rich in β-layers) in presence of the RNAs purified of their scrapie-associated fibrils. Analysis of the infectious recPrP-RNA mixture detected two populations of small RNAs of 27and 55 nucleotides in size, demonstrating a clear relation between the specific RNAs and the infectious properties of the prion amyloid (Simoneau et al., 2015).
Thus, there have been growing amount of proves of the role specific ncRNAs play in the pathogenesis of prion diseases. Since TEs is an important source of lncRNAs and micro RNAs, an assumption can be made that the individual com-position and specific changes of transposons in ontogenesis may be the element triggering the development of TSE and other cerebral proteinopathies including AD, PD and HD all characterized by a structural transformation of specific host proteins into new β-layer-rich conformations.
Conclusion
It has been assumed that interaction of a prion tertiary structure with certain gene sequences of lncRNAs may be one of the mechanisms of prion disease pathogenesis. Such interaction can be similar to the self-regulation of ncRNA by the products of its own translation. It has been found that ncRNAs can be translated, while the peptides formed has a regulatory effect on ncRNA genes. Association of microRNA polymorphism with prion disease development as well as changes in the expression of certain ncRNAs in TSE allow one to assume that specific ncRNAs are able to catalyze PrPC transformation into a PrPSc isoform. Therefore, optimal interactions of PrPSc with the genes of certain RNAs may cause the exponential growth of their amount, if the formed ncRNAs or the subproducts of their interaction are able to catalyze the transformation. MicroRNA activation by prion domains most likely potentiates the formation of microRNAs, whose regulation of the expression of certain genes may also stimulate the conformation transformation. It can also explain the species-specific character of prion infections as well as TSE long incubation period and fast progression. In TSE pathogenesis an important role is given to TEs to be the basis for the neural stem cell differentiation in the hippocampus triggered by TE ncRNAs.
Conflict of interest
The authors claim to have to conflict of interest to declare.
SARS-CoV-2 N protein, S protein and amyotrophic lateral sclerosis (ALS)
Also known as Lou Gehrig’s disease, ALS is a progressive nervous system disease that affects nerve cells in the brain and spinal cord, causing loss of muscle control. As this eventually affects control of the muscles needed to speak, eat and breathe it is a fatal disease, with no cure127.
Symptoms
Signs and symptoms of ALS vary greatly from person to person, depending on which neurons are affected. It generally begins with muscle weakness that spreads and gets worse over time. Signs and symptoms might include:
Difficulty walking or doing normal daily activities
Tripping and falling
Weakness in your legs, feet or ankles
Hand weakness or clumsiness
Slurred speech or trouble swallowing
Muscle cramps and twitching in your arms, shoulders and tongue
Inappropriate crying, laughing or yawning
Cognitive and behavioral changes
ALS often starts in the hands, feet or limbs, and then spreads to other parts of your body. As the disease advances and nerve cells are destroyed, your muscles get weaker. This eventually affects chewing, swallowing, speaking and breathing.
There's generally no pain in the early stages of ALS, and pain is uncommon in the later stages. ALS doesn't usually affect your bladder control or your senses.
Viral N-protein can impair the disassembly of stress granules (SGs) and promote ALS-associated amyloid aggregation. By Li et al (2022)128.
As discussed previously, FUS is one of the genes that encodes paraspeckle formation via the targeting of NEAT1:
Abstract
The nucleocapsid (N) protein of SARS-CoV-2 has been reported to have a high ability of liquid-liquid phase separation, which enables its incorporation into stress granules (SGs) of host cells. However, whether SG invasion by N protein occurs in the scenario of SARS-CoV-2 infection is unknow, neither do we know its consequence. Here, we used SARS-CoV-2 to infect mammalian cells and observed the incorporation of N protein into SGs, which resulted in markedly impaired self-disassembly but stimulated cell cellular clearance of SGs. NMR experiments further showed that N protein binds to the SG-related amyloid proteins via non-specific transient interactions, which not only expedites the phase transition of these proteins to aberrant amyloid aggregation in vitro, but also promotes the aggregation of FUS with ALS-associated P525L mutation in cells. In addition, we found that ACE2 is not necessary for the infection of SARS-CoV-2 to mammalian cells. Our work indicates that SARS-CoV-2 infection can impair the disassembly of host SGs and promote the aggregation of SG-related amyloid proteins, which may lead to an increased risk of neurodegeneration.
Several RNA-binding proteins of SGs including FUS, hnRNPA1, and TDP43 are meanwhile prone to undergo amyloid aggregation, which is closely associated with neurodegenerative diseases such as ALS and frontotemporal dementia (FTD).
N protein stimulates the aggregation of SG-related amyloid proteins
As we incubated N protein with FUS for 12 h, we observed that some LLPS droplets became spiky with fibrils growing out of the droplets (Fig. (Fig.3B3B and 3C). FRAP experiment showed that the fluorescence of these spiky droplets can hardly recover after bleaching, which confirms the solid nature of these droplets (Fig. (Fig.33B).
To further examine the influence of N protein on the amyloid fibril formation of FUS, hnRNPA1, and TDP43, we performed the ThT fluorescence assay and negative-staining transmission electron microscopy (TEM). Since the LC domains of FUS, hnRNPA1, and TDP43 are the amyloid-forming core sequences of these proteins (Johnson et al., 2009; Kato et al., 2012; Kim et al., 2013), we incubated N protein with the LC domains of these three SG proteins, respectively. The result showed that the presence of N protein markedly enhanced the ThT intensities of the fibril-forming samples, and shortened the lag time of the ThT kinetic curves in a dose-dependent manner (Fig. (Fig.5A).5A). TEM imaging confirmed amyloid fibril formation in these samples (Fig. (Fig.5B).5B). In addition, we confirmed that N protein under the examined conditions does not form amyloid fibrils (Fig. S8). These results indicate that N protein can generally stimulate the phase transition of the SG-related amyloid proteins into amyloid aggregation.
Figure 5.SARS-CoV-2 N protein promotes the amyloid aggregation of SG proteins. (A) ThT kinetic assays for amyloid fibril formation of the LC domains of FUS, TDP43, and hnRNPA1. Protein concentrations and molar ratios are indicated. Buffer for FUS-LC and hnRNPA1-LC amyloid formation: 50 mmol/L Tris-HCl, pH 7.5 and 100 mmol/L NaCl. Buffer for TDP43-LC amyloid formation: 50 mmol/L Bis-Tris, pH 6.5 and 100 mmol/L NaCl. Data correspond to mean ± SD, n = 3 independent replicates. (B) TEM images of samples in (A) at the end time point. Scale bar, 500 nm. (C) Confocal images of CFP-FUS P525L aggregation puncta in ACE2-HeLa cells infected by SARS-CoV-2. Cells without treatment of the virus are used as a control. FUS P525L is visualized by CFP fluorescence. N protein is immunostained with anti-N. The arrows indicate FUS P525L aggregates. Scale bar, 5 μm. Quantitative analysis of the aggregation area per cell in the imaging data is shown on the right. Values are means ± SD, n > 150 cells from 3 replicates. Student’s t-test
SARS-CoV-2 infection promotes amyloid aggregation of ALS-related FUS mutant in cells
We next sought to investigate SARS-CoV-2 infection on protein amyloid aggregation in cells. We used a cell model that overexpresses FUS with P525L mutation, a mutation found in ALS that disrupts the nuclear localization of FUS and results in FUS accumulation in the cytoplasm (Kwiatkowski et al., 2009; Dormann et al., 2010; De Santis et al., 2019). We first transfected ACE2-overexpressed HeLa cells with CFP-fused FUS P525L. Aggregation of FUS P525L in cells can be probed CFP fluorescence and an amyloid dye—pFTAA (Klingstedt et al., 2013; Qamar et al., 2018). Next, we treated the cells with SARS-CoV-2 and observed that the viral N protein colocalizes with FUS P525L aggregates (Fig. (Fig.5C).5C). Notably, comparing with the control cells (no virus infection), the aggregation of FUS P525L significantly increased upon virus infection (Fig. (Fig.5C).5C). We also observed the same enhancement of FUS P525L aggregation by using SARS-CoV-2 infected HeLa cells (Fig. S9A) and HeLa cells with overexpressed N protein (Fig. S9B). These data strengthen the potential consequence of SARS-CoV-2 infection in stimulating protein amyloid aggregation in the host cells.
Figure 6.Schematic diagram for the interplay between SARS-CoV-2 and host SGs. During SARS-CoV-2 replication in host cells, the viral N protein enters the host SGs and directly interacts with SG-related amyloid-forming proteins (e.g., FUS, hnRNPA1 and TDP43), which stimulates the liquid to solid phase transition (amyloid formation) of these host proteins. Inefficient maintenance of proteostasis may result in accumulation of the pathological amyloid fibrils and development of neurodegeneration
Our work provides molecular evidence for the increased risk of neurodegeneration after SARS-CoV-2 infection, and suggests paying a special attention to the incidence of neurodegenerative diseases in aged people under the current circumstances of ongoing widespread of SARS-CoV-2.
And from 2021, Idrees and Kumar published a paper on SARS-CoV-2 spike protein interactions with amyloidogenic proteins129. This doesn't exclude gp120 induced neurotoxicity130 or interactions with lncRNAs and miRNAs, as seen with HIV131:
Mechanisms involved up-regulation of hypoxia-inducible factor (HIF-1α), its translocation and binding to the long noncoding RNA (lncRNA) BACE1‐antisense transcript (BACE1-AS), resulting, in turn, in the formation of the BACE1-AS/BACE1 RNA complex, subsequently leading to increased BACE1 protein, and activity and generation of Aβ-42.
Abstract
The post-infection of COVID-19 includes a myriad of neurologic symptoms including neurodegeneration. Protein aggregation in brain can be considered as one of the important reasons behind the neurodegeneration. SARS-CoV-2 Spike S1 protein receptor binding domain (SARS-CoV-2 S1 RBD) binds to heparin and heparin binding proteins. Moreover, heparin binding accelerates the aggregation of the pathological amyloid proteins present in the brain. In this paper, we have shown that the SARS-CoV-2 S1 RBD binds to a number of aggregation-prone, heparin binding proteins including Aβ, α-synuclein, tau, prion, and TDP-43 RRM. These interactions suggests that the heparin-binding site on the S1 protein might assist the binding of amyloid proteins to the viral surface and thus could initiate aggregation of these proteins and finally leads to neurodegeneration in brain. The results will help us to prevent future outcomes of neurodegeneration by targeting this binding and aggregation process.
Keywords: COVID-19, Heparin, Heparin binding proteins, Neurodegeneration, Protein aggregation, SARS-CoV-2
Very recently, Tavassoly et al. proposed a view that seeded protein aggregation by SARS-CoV-2 could be attributed to long-term post-infection complications including neurodegeneration [4]. They suggested that SARS-CoV-2 spike protein S1 region binds to heparin and heparin binding proteins (HBPs) present in brain which are prone to self-assembly, aggregation, and fibrillation processes. They also showed that the peptide from S protein (S–CoV-peptide; ∼150 aa) has more aggregation formation propensity than the known aggregation-prone proteins, suggesting that this peptide is prone to act as functional amyloid and form toxic aggregates. Thus, the heparin binding and aggregation propensity of S1 protein has been suggested the ability of S1 to form amyloid and toxic aggregates that can act as seeds to aggregate many of the misfolded brain proteins and can ultimately leads to neurodegeneration. It has been suggested that SARS-CoV-2 infection invades the CNS by controlling protein synthesis machinery, disturbs endoplasmic reticulum and mitochondrial function and increases the accumulation of misfolded proteins, thereby activates protein aggregation, mitochondrial oxidative stress, apoptosis and neurodegeneration [3,5,10].
COVID‐19–accelerated disease progression in two patients with pre-existing ALS by Li and Bedlack (2021)132. These case reports support the chain reaction hypothesis:
We report two patients with slowly progressive amyotrophic lateral sclerosis (ALS) who experienced rapid functional decline after contracting severe acute respiratory virus–coronavirus‐2 (SARS‐CoV‐2).
A 78‐year‐old man developed proximal leg weakness in 2014 and was diagnosed with probable ALS based on El Escorial criteria1 in 2016. His revised ALS Functional Rating Scale (ALSFRS‐R) assessment, administered by physicians, showed scoring that progressed at an unusually slow rate since that time (Figure 1A). In December 2020, the patient developed a fever, and SARS‐CoV‐2 viral polymerase chain reaction of the nasopharyngeal swab was positive. One week later, he presented to an emergency department with fever, shortness of breath, and delirium. Computed tomography scan of the chest revealed signs of pneumonia. He also developed deep venous thrombosis in the left femoral vein but did not have a pulmonary embolism. The patient was hospitalized and treated with dexamethasone, remdesivir, oxygen supplement, and anticoagulation. He did not require mechanical ventilation. The patient was discharged 5 days later when he became afebrile and his peripheral capillary oxygen saturation (SpO2) was 93% on room air. Despite prompt treatment and recovery from pneumonia, his functional status declined rapidly. His ALSFRS‐R score that the physician administered during follow‐up telemedicine visits decreased by 16 points within 4 months, 11 points in the final month before he died of hypercarbic respiratory failure.
FIGURE 1. Rapid decline of revised ALSFRS‐R in two amyotrophic lateral sclerosis patients post–COVID‐19 infection. A, Slow decline of ALSFRS‐R in patient 1, which plateaued from January 2019 to September 2020, but declined rapidly after COVID‐19 infection. B, Stable ALSFRS‐R between 2013 and 2019 in patient 2, followed by a rapid decline in 2020 after COVID‐19 infection. ↓ indicates postinfection ALSFRS‐R. Abbreviation: ALSFRS‐R, revised Amyotropic Lateral Sclerosis Functional Rating Scale
A 66‐year‐old man developed his first sign of weakness in his legs in 2004 and was diagnosed with laboratory‐supported probable ALS in 2006. His disease had an unusually slow progression. Between 2014 and 2019, the ALSFRS‐R, administered by physicians, remained stable at 36 to 38. He had bilateral foot dorsiflexion and plantarflexion weakness as well as mild hand weakness during his clinic visit in December 2019. His bulbar and respiratory function were both normal. In April 2020, he contracted the virus after several family members had COVID‐19 infection. His infection was confirmed by a positive SARS‐CoV‐2 viral polymerase chain reaction of a nasopharyngeal swab sample. His symptoms included a low‐grade fever for 1 day, and fatigue, loss of smell, and loss of taste for 1 week. He was never hospitalized. After the COVID infection, he noticed fast progression of arm and leg weakness. He is now wheelchair‐dependent.He also developed mild‐to‐moderate speech and swallowing difficulty 4 months after the COVID‐19 infection. ALSFRS‐R score administered during a telemedicine visit decreased by 5 points when compared with December 2019 (Figure 1B).
Structure-function relationships of lncRNA mechanism
It is astounding to me that as recently as 2020 the 3D structure and functional relationships of select lncRNAs with experimental gene therapy agents were unknown before proceeding to inject them into 4 billion arms. M1 macrophage skewing and dysregulated interferon responses being just one consequence of this lack of research.
Indeed, Karissa Sanbonmatsu called the prospect of drugging lncRNAs “daunting” without this information133. And, as we have seen, drugging lncRNAs is exactly what spike protein mRNA does - its the very definition of a "protein based drug":
While long non-coding RNAs are known to play key roles in disease and development, relatively few structural studies have been performed for this important class of RNAs. Here, we review functional studies of long non-coding RNAs and expose the need for high-resolution 3-D structural studies, discussing the roles of long non-coding RNAs in the cell and how structure–function relationships might be used to elucidate further understanding.
An ironic reference to COVID-19 vaccine development:
Because this class of RNA molecules has been found to be important for processes in cancer, development, and brain function, there is keen interest in the pharmaceutical community (Kashi et al. 1859; Hon et al. 2017). However, the enormous size of these RNAs, which are often kilobases or tens of kilobases in length, makes the prospects of drugging them daunting. If the drug is a small molecule, then which of the 10,000 bases on a 10 kb lncRNA should be targeted? If the drug is an antisense oligo, which region of the RNA should be targeted? Which regions should not be targeted?
The steps of pre-clinical trials, clinical trials, and regulatory approval have been in the news lately regarding COVID-19 vaccine development. Similar steps are required for protein-based drugs, such as anti-viral therapeutics, cancer drugs, anti-depressants, antibiotics, and disease-related therapies (Matthews et al. 2016). However, before these steps can begin, target identification, lead generation, lead optimization, and drug candidate selection must take place. Each of these stages requires considerable structural characterization. In the case of protein-based drugs, often a high-resolution 3-D structure of the target protein is solved by either X-ray crystallography or cryo-EM, followed by binding pocket characterization, hit identification, lead development, and lead optimization (Grey and Thompson 2010). Currently, there are no high-resolution 3-D structures of lncRNAs.
Relative to the history of structural biology in mechanistic studies and drug development in the protein community, and the fact that high-resolution structures of lncRNAs have not yet been solved, lncRNA mechanism is not well understood at the molecular level of detail. Without a clear understanding of structure, structure–function relationships, and mechanism, lncRNA drug discovery is in its early stages. In the case of lncRNAs, we expect understanding mechanism will require determination of the structure–function relationship for the RNA, and determination of the structure–function relationship will require solving the lncRNA 3-D structure at high resolution, similar to how structure–function relationships and mechanisms were worked out for proteins. Thus, we anticipate that solving structures of lncRNAs will be an important stage for determination of lncRNA mechanism and for lncRNA drug discovery.
In biological systems, we often first know that a molecule is important, and even what it does, but not how it does it. The ‘how,’ in the case of a protein, is then worked out by solving the protein’s 3-D structure and relating it to its function.
Once we know the ‘how,’ we can begin to understand the molecule in context and start thinking about drugging the molecule. In the case of protein molecules, their function almost always hinges on interaction with another molecule, such as another protein, RNA, or DNA molecule.Solving the structure of the protein in isolation and complexed with its target molecules produces invaluable information about its function and about the structure–function relationship.
Therapeutics
This subject requires a further Substack, but a search of PubMed seems to indicate that more research has been conducted into the effects of natural or repurposed therapeutics on lncRNAs than has been conducted on mass distributed mRNA gene therapy agents, or at least that we have been allowed to see.
Examples include baicalein and cancer; quercetin and cancer prevention; berberine and coronary heart disease; artemisinin and atherosclerosis; icariin and osteoarthritis; silymarin and colon cancer; metformin and breast cancer; resveratrol and the inhibition of cancer cell proliferation.
As is often the case we can thank Chinese researchers for conducting almost all of this research, western research is notably absent.
On balance, a milder infection was the worst case scenario for cancer patients as MALAT1 was overexpressed in the CD4+ T cells of patients with mild infections.
More research is needed to confirm the interactions between human lncRNAs and spike mRNA (+ viral vs synthetic), and to screen for potential RNA–RNA, RNA–DNA, and RNA–protein complexes and the pathologies associated with this, such as cardiovascular disease, immunosuppression, autoimmune disorders or oncogenesis and metastasis.
3D structural analysis of key lncRNAs also needs conducting so that potential pathological interactions via binding sites can be predicted in silico.
Additionally, all the currently administered and future vaccines, including for delivery at point of care, should be screened for potentially pathological effects vis dysregulated expression of lncRNAs and for any long term immunoglobulin subclass changes, as discussed in the last Substack.
These experimental gene therapy mRNA products should be withdrawn immediately, especially from cancer patients as per the precautionary principle.
Notwithstanding the foregoing, we already have a multitude of safety and efficacy signals raised since the early trial stages which were fraudulently ignored or misclassified as unrelated.
DoorlessCarp’s Scientific Literature Reviews is a reader-supported publication. To receive new posts and support my work, consider becoming a free or paid subscriber. Thank you all for your kind support, it would not be possible to sustain this research without it.
NUCLEUS TO RIBOSOMES TO ER GIF FROM NPR: PROTEIN SYNTHESIS
Sebastian-delaCruz M, Gonzalez-Moro I, Olazagoitia-Garmendia A, Castellanos-Rubio A, Santin I. The Role of lncRNAs in Gene Expression Regulation through mRNA Stabilization.Non-Coding RNA. 2021; 7(1):3. https://doi.org/10.3390/ncrna7010003
Lukiw WJ, Handley P, Wong L, Crapper McLachlan DR (June 1992). "BC200 RNA in normal human neocortex, non-Alzheimer dementia (NAD), and senile dementia of the Alzheimer type (AD)". Neurochemical Research. 17 (6): 591–597. doi:10.1007/bf00968788. PMID1603265. S2CID39305428.
Fu X, Ravindranath L, Tran N, Petrovics G, Srivastava S (March 2006). "Regulation of apoptosis by a prostate-specific and prostate cancer-associated noncoding gene, PCGEM1". DNA and Cell Biology. 25 (3): 135–141. doi:10.1089/dna.2006.25.135. PMID16569192.
Boeckel JN, Perret MF, Glaser SF, Seeger T, Heumüller AW, Chen W, et al. (January 2019). "Identification and regulation of the long non-coding RNA Heat2 in heart failure". Journal of Molecular and Cellular Cardiology. 126: 13–22. doi:10.1016/j.yjmcc.2018.11.004. PMID30445017. S2CID53569324.
Yi C, Zhao Z, Wang S, Sun X, Zhang D, Sun X, Zhang A, Jin M. Influenza A Virus PA Antagonizes Interferon-β by Interacting with Interferon Regulatory Factor 3. Front Immunol. 2017 Sep 11;8:1051. doi: 10.3389/fimmu.2017.01051. PMID: 28955326; PMCID: PMC5600993.
Yang Y, Zhang Z, Wu Z, Lin W, Yu M. Downregulation of the expression of the lncRNA MIAT inhibits melanoma migration and invasion through the PI3K/AKT signaling pathway. Cancer Biomark. 2019;24(2):203-211. doi: 10.3233/CBM-181869. PMID: 30614798.
James P. Hewitson, Katie A. West, Kylie R. James, Gulab Fatima Rani, Nidhi Dey, Audrey Romano, Najmeeyah Brown, Sarah A. Teichmann, Paul M. Kaye, Dimitris Lagos; Malat1 Suppresses Immunity to Infection through Promoting Expression of Maf and IL-10 in Th Cells. J Immunol 1 June 2020; 204 (11): 2949–2960. https://doi.org/10.4049/jimmunol.1900940
Islam H, Chamberlain TC, Mui AL, Little JP. Elevated Interleukin-10 Levels in COVID-19: Potentiation of Pro-Inflammatory Responses or Impaired Anti-Inflammatory Action? Front Immunol. 2021 Jun 21;12:677008. doi: 10.3389/fimmu.2021.677008. PMID: 34234779; PMCID: PMC8255680.
Qu D, Sun WW, Li L, Ma L, Sun L, Jin X, Li T, Hou W, Wang JH. Long noncoding RNA MALAT1 releases epigenetic silencing of HIV-1 replication by displacing the polycomb repressive complex 2 from binding to the LTR promoter. Nucleic Acids Res. 2019 Apr 8;47(6):3013-3027. doi: 10.1093/nar/gkz117. PMID: 30788509; PMCID: PMC6451131.
Ruiz Ramírez, A. V., & Prado Montes de Oca, E. (2022). Therapeutic Potential of Long Non-Coding RNAs of HIV-1, SARS-CoV-2, and Endogenous Retroviruses.Frontiers in Virology, 2. https://www.frontiersin.org/articles/10.3389/fviro.2022.849349
Fan Y, Sanyal S, Bruzzone R. Breaking Bad: How Viruses Subvert the Cell Cycle. Front Cell Infect Microbiol. 2018 Nov 19;8:396. doi: 10.3389/fcimb.2018.00396. PMID: 30510918; PMCID: PMC6252338.
Aloni-Grinstein R, Charni-Natan M, Solomon H, Rotter V. p53 and the Viral Connection: Back into the Future‡. Cancers (Basel). 2018 Jun 4;10(6):178. doi: 10.3390/cancers10060178. PMID: 29866997; PMCID: PMC6024945.
Kanduc D. From Anti-Severe Acute Respiratory Syndrome Coronavirus 2 Immune Response to Cancer Onset via Molecular Mimicry and Cross-Reactivity. Glob Med Genet. 2021 Sep 7;8(4):176-182. doi: 10.1055/s-0041-1735590. PMID: 34877576; PMCID: PMC8635832.
Peeters M, Verbruggen L, Teuwen L, Vanhoutte G, Vande Kerckhove S, Peeters B, Raats S, Van der Massen I, De Keersmaecker S, Debie Y, Huizing M, Pannus P, Neven K, Ariën KK, Martens GA, Van Den Bulcke M, Roelant E, Desombere I, Anguille S, Goossens M, Vandamme T, van Dam P. Reduced humoral immune response after BNT162b2 coronavirus disease 2019 messenger RNA vaccination in cancer patients under antineoplastic treatment. ESMO Open. 2021 Oct;6(5):100274. doi: 10.1016/j.esmoop.2021.100274. Epub 2021 Sep 8. PMID: 34597941; PMCID: PMC8423808.
Landskron G, De la Fuente M, Thuwajit P, Thuwajit C, Hermoso MA. Chronic inflammation and cytokines in the tumor microenvironment. J Immunol Res. 2014;2014:149185. doi: 10.1155/2014/149185. Epub 2014 May 13. PMID: 24901008; PMCID: PMC4036716.
Tang, M., Hu, X., Wang, Y., Yao, X., Zhang, W., Yu, C., Cheng, F., Li, J., & Fang, Q. (2021). Ivermectin, a potential anticancer drug derived from an antiparasitic drug. In Pharmacological Research (Vol. 163). https://doi.org/10.1016/j.phrs.2020.105207
Zhou S, Wu H, Ning W, Wu X, Xu X, Ma Y, Li X, Hu J, Wang C, Wang J. Ivermectin has New Application in Inhibiting Colorectal Cancer Cell Growth. Front Pharmacol. 2021 Aug 13;12:717529. doi: 10.3389/fphar.2021.717529. PMID: 34483925; PMCID: PMC8415024.
Jullien M, Le Bourgeois A, Coste-Burel M, Peterlin P, Garnier A, Audrain M, Rimbert M, Imbert BM, Drumel T, Le Gouill S, Moreau P, Mahé B, Dubruille V, Blin N, Lok A, Touzeau C, Gastinne T, Tessoulin B, Vantyghem S, Béné MC, Guillaume T, Chevallier P. Profound B-Cell Lymphopenia Is a Major Factor Predicting Poor Humoral Response after BNT162b2 mRNA Sars-Cov-2 Vaccines in Recipients of Allogeneic Hematopoietic Stem Cell Transplantation. Blood. 2021 Nov 23;138:3911. doi: 10.1182/blood-2021-150439. Epub 2021 Dec 24. PMCID: PMC8701579.
Fujii, Y. R. (2021). Quantum microRNA Assessment of COVID-19 RNA Vaccine: Hidden Potency of BNT162b2 SASR-CoV-2 Spike RNA as MicroRNA Vaccine.Advancements in Case Studies, 3(1). https://doi.org/10.31031/aics.2021.03.000552
Huang K, Wang C, Vagts C, Raguveer V, Finn PW, Perkins DL. Long non-coding RNAs (lncRNAs) NEAT1 and MALAT1 are differentially expressed in severe COVID-19 patients: An integrated single-cell analysis. PLoS One. 2022 Jan 10;17(1):e0261242. doi: 10.1371/journal.pone.0261242. PMID: 35007307; PMCID: PMC8746747.
Masters, S. L., Gerlic, M., Metcalf, D., Preston, S. P., Pellegrini, M., O’Donnell, J. A., McArthur, K., Baldwin, T. M., Chevrier, S., Nowell, C. J., Cengia, L. H., Henley, K. J., Collinge, J. E., Kastner, D. L., Feigenbaum, L., Hilton, D. J., Alexander, W. S., Croker, B. A., & Kile, B. T. (2012). Activation of the NLRP1 Inflammasome Induces the Pyroptotic Death of Hematopoietic Progenitor Cells.Blood, 120(21). https://doi.org/10.1182/blood.v120.21.1213.1213
Mills CD. M1 and M2 Macrophages: Oracles of Health and Disease. Crit Rev Immunol. 2012;32(6):463-88. doi: 10.1615/critrevimmunol.v32.i6.10. PMID: 23428224.
Saha C, Laha S, Chatterjee R, Bhattacharyya NP. Co-Regulation of Protein Coding Genes by Transcription Factor and Long Non-Coding RNA in SARS-CoV-2 Infected Cells: An In Silico Analysis. Noncoding RNA. 2021 Nov 29;7(4):74. doi: 10.3390/ncrna7040074. PMID: 34940755; PMCID: PMC8708613.
Pisani G, Baron B. NEAT1 and Paraspeckles in Cancer Development and Chemoresistance. Noncoding RNA. 2020 Oct 30;6(4):43. doi: 10.3390/ncrna6040043. PMID: 33143162; PMCID: PMC7712271.
Xiong, M., Wu, M., Dan Peng et al.LncRNA DANCR represses Doxorubicin-induced apoptosis through stabilizing MALAT1 expression in colorectal cancer cells.Cell Death Dis12, 24 (2021). https://doi.org/10.1038/s41419-020-03318-8
Yunqi Lu, Zhongyi Hu, Lingegowda S. Mangala, Zachary E. Stine, Xiaowen Hu, Dahai Jiang, Yan Xiang, Youyou Zhang, Sunila Pradeep, Cristian Rodriguez-Aguayo, Gabriel Lopez-Berestein, Angelo M. DeMarzo, Anil K. Sood, Lin Zhang, Chi V. Dang; MYC Targeted Long Noncoding RNA DANCR Promotes Cancer in Part by Reducing p21 Levels.Cancer Res 1 January 2018; 78 (1): 64–74. https://doi.org/10.1158/0008-5472.CAN-17-0815
Greco A. Involvement of the nucleolus in replication of human viruses. Rev Med Virol. 2009 Jul;19(4):201-14. doi: 10.1002/rmv.614. PMID: 19399920; PMCID: PMC7169183.
Laha S, Saha C, Dutta S, Basu M, Chatterjee R, Ghosh S, Bhattacharyya NP. In silico analysis of altered expression of long non-coding RNA in SARS-CoV-2 infected cells and their possible regulation by STAT1, STAT3 and interferon regulatory factors. Heliyon. 2021 Mar;7(3):e06395. doi: 10.1016/j.heliyon.2021.e06395. Epub 2021 Feb 27. PMID: 33688586; PMCID: PMC7914022.
Yang Y, Li M, Ma Y, Ye W, Si Y, Zheng X, Liu H, Cheng L, Zhang L, Zhang H, Zhang X, Lei Y, Shen L, Zhang F, Ma H. LncRNA NEAT1 Potentiates SREBP2 Activity to Promote Inflammatory Macrophage Activation and Limit Hantaan Virus Propagation. Front Microbiol. 2022 Apr 13;13:849020. doi: 10.3389/fmicb.2022.849020. PMID: 35495674; PMCID: PMC9044491.
Meydan C, Madrer N, Soreq H. The Neat Dance of COVID-19: NEAT1, DANCR, and Co-Modulated Cholinergic RNAs Link to Inflammation. Front Immunol. 2020 Oct 9;11:590870. doi: 10.3389/fimmu.2020.590870. PMID: 33163005; PMCID: PMC7581732.
Moazzam-Jazi M, Lanjanian H, Maleknia S, Hedayati M, Daneshpour MS. Interplay between SARS-CoV-2 and human long non-coding RNAs. J Cell Mol Med. 2021 Jun;25(12):5823-5827. doi: 10.1111/jcmm.16596. Epub 2021 May 9. PMID: 33969601; PMCID: PMC8184717.
Natarelli L, Parca L, Mazza T, Weber C, Virgili F, Fratantonio D. MicroRNAs and Long Non-Coding RNAs as Potential Candidates to Target Specific Motifs of SARS-CoV-2. Noncoding RNA. 2021 Feb 18;7(1):14. doi: 10.3390/ncrna7010014. PMID: 33670580; PMCID: PMC7931055.
Zhong Y, Ashley CL, Steain M, Ataide SF. Assessing the suitability of long non-coding RNAs as therapeutic targets and biomarkers in SARS-CoV-2 infection. Front Mol Biosci. 2022 Aug 16;9:975322. doi: 10.3389/fmolb.2022.975322. PMID: 36052163; PMCID: PMC9424846.
Jiang W, Zhao S, Jiang X, Zhang E, Hu G, Hu B, Zheng P, Xiao J, Lu Z, Lu Y, Ni J, Chen C, Wang X, Yang L, Wan R. The circadian clock gene Bmal1 acts as a potential anti-oncogene in pancreatic cancer by activating the p53 tumor suppressor pathway. Cancer Lett. 2016 Feb 28;371(2):314-25. doi: 10.1016/j.canlet.2015.12.002. Epub 2015 Dec 9. PMID: 26683776.
Kallen AN, Zhou XB, Xu J, Qiao C, Ma J, Yan L, Lu L, Liu C, Yi JS, Zhang H, Min W, Bennett AM, Gregory RI, Ding Y, Huang Y. The imprinted H19 lncRNA antagonizes let-7 microRNAs. Mol Cell. 2013 Oct 10;52(1):101-12. doi: 10.1016/j.molcel.2013.08.027. Epub 2013 Sep 19. PMID: 24055342; PMCID: PMC3843377.
Pontecorvi G, Bellenghi M, Ortona E, Carè A. microRNAs as new possible actors in gender disparities of Covid-19 pandemic. Acta Physiol (Oxf). 2020 Sep;230(1):e13538. doi: 10.1111/apha.13538. Epub 2020 Jul 30. PMID: 32672403; PMCID: PMC7404333.
Ren M, Wang T, Wei X, Wang Y, Ouyang C, Xie Y, Ye X, Han Z. LncRNA H19 regulates smooth muscle cell functions and participates in the development of aortic dissection through sponging miR-193b-3p. Biosci Rep. 2021 Jan 29;41(1):BSR20202298. doi: 10.1042/BSR20202298. PMID: 33403385; PMCID: PMC7823186.
Zappa M, Verdecchia P, Spanevello A, Angeli F. Structural evolution of severe acute respiratory syndrome coronavirus 2: Implications for adhesivity to angiotensin-converting enzyme 2 receptors and vaccines. Eur J Intern Med. 2022 Oct;104:33-36. doi: 10.1016/j.ejim.2022.08.012. Epub 2022 Aug 12. PMID: 35985948; PMCID: PMC9372025.
Renna NF, de Las Heras N, Miatello RM. Pathophysiology of vascular remodeling in hypertension. Int J Hypertens. 2013;2013:808353. doi: 10.1155/2013/808353. Epub 2013 Jul 22. PMID: 23970958; PMCID: PMC3736482.
Xu TP, Huang MD, Xia R, Liu XX, Sun M, Yin L, Chen WM, Han L, Zhang EB, Kong R, De W, Shu YQ. Decreased expression of the long non-coding RNA FENDRR is associated with poor prognosis in gastric cancer and FENDRR regulates gastric cancer cell metastasis by affecting fibronectin1 expression. J Hematol Oncol. 2014 Aug 29;7:63. doi: 10.1186/s13045-014-0063-7. PMID: 25167886; PMCID: PMC4237812.
Wang J, Cai Y, Lu H, Zhang F, Zheng J. LncRNA APOA1-AS facilitates proliferation and migration and represses apoptosis of VSMCs through TAF15-mediated SMAD3 mRNA stabilization. Cell Cycle. 2021 Sep;20(17):1642-1652. doi: 10.1080/15384101.2021.1951940. Epub 2021 Aug 12. PMID: 34382908; PMCID: PMC8489913.
Ye M, Ma J, Liu B, Liu X, Ma D, Dong K. Linc01105 acts as an oncogene in the development of neuroblastoma. Oncol Rep. 2019 Oct;42(4):1527-1538. doi: 10.3892/or.2019.7257. Epub 2019 Aug 2. Erratum in: Oncol Rep. 2021 Sep;46(3): PMID: 31524279.
Cheng MH, Zhang S, Porritt RA, Noval Rivas M, Paschold L, Willscher E, Binder M, Arditi M, Bahar I. Superantigenic character of an insert unique to SARS-CoV-2 spike supported by skewed TCR repertoire in patients with hyperinflammation. Proc Natl Acad Sci U S A. 2020 Oct 13;117(41):25254-25262. doi: 10.1073/pnas.2010722117. Epub 2020 Sep 28. PMID: 32989130; PMCID: PMC7568239.
Harjunpää, H., Asens, M. L., Guenther, C., & Fagerholm, S. C. (2019). Cell adhesion molecules and their roles and regulation in the immune and tumor microenvironment. In Frontiers in Immunology (Vol. 10, Issue MAY). https://doi.org/10.3389/fimmu.2019.01078
Schimmel L, Heemskerk N, van Buul JD. Leukocyte transendothelial migration: A local affair. Small GTPases. 2017 Jan 2;8(1):1-15. doi: 10.1080/21541248.2016.1197872. Epub 2016 Aug 15. PMID: 27715453; PMCID: PMC5331897.
Yoder, A., Yu, D., Dong, L., Iyer, S. R., Xu, X., Kelly, J., Liu, J., Wang, W., Vorster, P. J., Agulto, L., Stephany, D. A., Cooper, J. N., Marsh, J. W., & Wu, Y. (2008). HIV Envelope-CXCR4 Signaling Activates Cofilin to Overcome Cortical Actin Restriction in Resting CD4 T Cells. <i>Cell</i>, <i>134</i>(5). https://doi.org/10.1016/j.cell.2008.06.036</div>
de Iongh RU, Wederell E, Lovicu FJ, McAvoy JW. Transforming growth factor-beta-induced epithelial-mesenchymal transition in the lens: a model for cataract formation. Cells Tissues Organs. 2005;179(1-2):43-55. doi: 10.1159/000084508. PMID: 15942192.
Mariotti, B., Servaas, N. H., Rossato, M., Tamassia, N., Cassatella, M. A., Cossu, M., Beretta, L., van der Kroef, M., Radstake, T. R. D. J., & Bazzoni, F. (2019). The long non-coding RNA NRIR drives IFN-Response in monocytes: Implication for systemic sclerosis. <i>Frontiers in Immunology</i>, <i>10</i>(JAN). https://doi.org/10.3389/fimmu.2019.00100</div>
Joachims ML, Khatri B, Li C, et al. Dysregulated long non-coding RNA in Sjögren’s disease impacts both interferon and adaptive immune responses. RMD Open 2022;8:e002672. doi: 10.1136/rmdopen-2022-002672
Barriocanal M, Carnero E, Segura V, Fortes P. Long Non-Coding RNA BST2/BISPR is Induced by IFN and Regulates the Expression of the Antiviral Factor Tetherin. Front Immunol. 2015 Jan 9;5:655. doi: 10.3389/fimmu.2014.00655. PMID: 25620967; PMCID: PMC4288319.
Guruswamy Mahesh and Roopa Biswas. MicroRNA-155: A Master Regulator of Inflammation. Journal of Interferon & Cytokine Research. Jun 2019.321-330.http://doi.org/10.1089/jir.2018.0155
Liu L, Wu Y, Chen W, Li Y, Yu J, Zhang G, Fu P, Huang L, Xiong J, Jie Z. The m7G-Related Long Noncoding RNA Signature Predicts Prognosis and Indicates Tumour Immune Infiltration in Colon Cancer. Front Genet. 2022 Jun 29;13:892589. doi: 10.3389/fgene.2022.892589. PMID: 35846130; PMCID: PMC9277109.
Polansky H, Schwab H. How latent viruses cause breast cancer: An explanation based on the microcompetition model. Bosn J Basic Med Sci. 2019 Aug 20;19(3):221-226. doi: 10.17305/bjbms.2018.3950. PMID: 30579323; PMCID: PMC6716096.
Zhang P, Lu B, Zhang Q, Xu F, Zhang R, Wang C, Liu Y, Wei C, Mei L. LncRNA NEAT1 Sponges MiRNA-148a-3p to Suppress Choroidal Neovascularization and M2 macrophage polarization. Mol Immunol. 2020 Nov;127:212-222. doi: 10.1016/j.molimm.2020.08.008. Epub 2020 Oct 1. PMID: 33011404.
Huang F, Zhao JL, Wang L, Gao CC, Liang SQ, An DJ, Bai J, Chen Y, Han H, Qin HY. miR-148a-3p Mediates Notch Signaling to Promote the Differentiation and M1 Activation of Macrophages. Front Immunol. 2017 Oct 16;8:1327. doi: 10.3389/fimmu.2017.01327. PMID: 29085372; PMCID: PMC5650608.
Janghorban M, Xin L, Rosen JM, Zhang XH. Notch Signaling as a Regulator of the Tumor Immune Response: To Target or Not To Target? Front Immunol. 2018 Jul 16;9:1649. doi: 10.3389/fimmu.2018.01649. PMID: 30061899; PMCID: PMC6055003.
Javier J, Hinge A, Bartram J, Xu J, Filippi MD. Transforming growth factor-β signaling modifies the hematopoietic acute inflammatory response to drive bone marrow failure. Haematologica. 2022 Jun 1;107(6):1323-1334. doi: 10.3324/haematol.2020.273292. PMID: 34706493; PMCID: PMC9152956.
Liu Z, Huang XR, Chen HY, Penninger JM, Lan HY. Loss of angiotensin-converting enzyme 2 enhances TGF-β/Smad-mediated renal fibrosis and NF-κB-driven renal inflammation in a mouse model of obstructive nephropathy. Lab Invest. 2012 May;92(5):650-61. doi: 10.1038/labinvest.2012.2. Epub 2012 Feb 13. PMID: 22330342.
Casamassima F, Ruggiero C, Caramella D, Tinacci E, Villari N, Ruggiero M. Hematopoietic bone marrow recovery after radiation therapy: MRI evaluation. Blood. 1989 May 1;73(6):1677-81. PMID: 2713500.
Liu L, Liu F, Sun Z, Peng Z, You T, Yu Z. LncRNA NEAT1 promotes apoptosis and inflammation in LPS-induced sepsis models by targeting miR-590-3p. Exp Ther Med. 2020 Oct;20(4):3290-3300. doi: 10.3892/etm.2020.9079. Epub 2020 Jul 29. PMID: 32855700; PMCID: PMC7444425.
Zhao H, Wang Y, Hou W, Ding X, Wang W. Long non-coding RNA MALAT1 promotes cell proliferation, migration and invasion by targeting miR-590-3p in osteosarcoma. Exp Ther Med. 2022 Sep 14;24(5):672. doi: 10.3892/etm.2022.11608. PMID: 36277152; PMCID: PMC9533361.
Chattopadhyay P, Mishra P, Mehta P, Soni J, Gupta R, Tarai B, Budhiraja S, Pandey R. Transcriptomic study reveals lncRNA-mediated downregulation of innate immune and inflammatory response in the SARS-CoV-2 vaccination breakthrough infections. Front Immunol. 2022 Nov 18;13:1035111. doi: 10.3389/fimmu.2022.1035111. PMID: 36466827; PMCID: PMC9716354.
Rodić N, Sharma R, Sharma R, Zampella J, Dai L, Taylor MS, Hruban RH, Iacobuzio-Donahue CA, Maitra A, Torbenson MS, Goggins M, Shih IeM, Duffield AS, Montgomery EA, Gabrielson E, Netto GJ, Lotan TL, De Marzo AM, Westra W, Binder ZA, Orr BA, Gallia GL, Eberhart CG, Boeke JD, Harris CR, Burns KH. Long interspersed element-1 protein expression is a hallmark of many human cancers. Am J Pathol. 2014 May;184(5):1280-6. doi: 10.1016/j.ajpath.2014.01.007. Epub 2014 Mar 6. PMID: 24607009; PMCID: PMC4005969.
Lee KW, Pausova Z. Cigarette smoking and DNA methylation. Front Genet. 2013 Jul 17;4:132. doi: 10.3389/fgene.2013.00132. PMID: 23882278; PMCID: PMC3713237.
Mushegian AR, Elena SF. RNAs That Behave Like Prions. mSphere. 2020 Jul 1;5(4):e00520-20. doi: 10.1128/mSphere.00520-20. PMID: 32611703; PMCID: PMC7333576.
Mustafin, R. N., & KhusnutDinova, E. K. (2018). Interrelation of prions with non-coDing RNAs. In Vavilovskii Zhurnal Genetiki i Selektsii (Vol. 22, Issue 4). https://doi.org/10.18699/VJ18.377
Li Y, Lu S, Gu J, Xia W, Zhang S, Zhang S, Wang Y, Zhang C, Sun Y, Lei J, Liu C, Su Z, Yang J, Peng X, Li D. SARS-CoV-2 impairs the disassembly of stress granules and promotes ALS-associated amyloid aggregation. Protein Cell. 2022 Aug;13(8):602-614. doi: 10.1007/s13238-022-00905-7. Epub 2022 Apr 6. PMID: 35384603; PMCID: PMC8983322.
Idrees D, Kumar V. SARS-CoV-2 spike protein interactions with amyloidogenic proteins: Potential clues to neurodegeneration. Biochem Biophys Res Commun. 2021 May 21;554:94-98. doi: 10.1016/j.bbrc.2021.03.100. Epub 2021 Mar 24. PMID: 33789211; PMCID: PMC7988450.
Louboutin JP, Agrawal L, Reyes BA, Van Bockstaele EJ, Strayer DS. HIV-1 gp120 neurotoxicity proximally and at a distance from the point of exposure: protection by rSV40 delivery of antioxidant enzymes. Neurobiol Dis. 2009 Jun;34(3):462-76. doi: 10.1016/j.nbd.2009.03.003. Epub 2009 Mar 25. PMID: 19327399.
Li X, Bedlack R. COVID-19-accelerated disease progression in two patients with amyotrophic lateral sclerosis. Muscle Nerve. 2021 Sep;64(3):E13-E15. doi: 10.1002/mus.27351. Epub 2021 Jun 23. PMID: 34131925; PMCID: PMC8441768.
Sanbonmatsu K. Getting to the bottom of lncRNA mechanism: structure-function relationships. Mamm Genome. 2022 Jun;33(2):343-353. doi: 10.1007/s00335-021-09924-x. Epub 2021 Oct 12. PMID: 34642784; PMCID: PMC8509902.
Well, I’m 17% done, and bookmarked. This will have to be achieved in 3-5 sessions, considering my ‘Layman’ and ‘ESL (English was actually my fourth not second language, when I was already a in high school unfortunately)’ statuses.
Nonetheless, thank you, and very impressive so far.








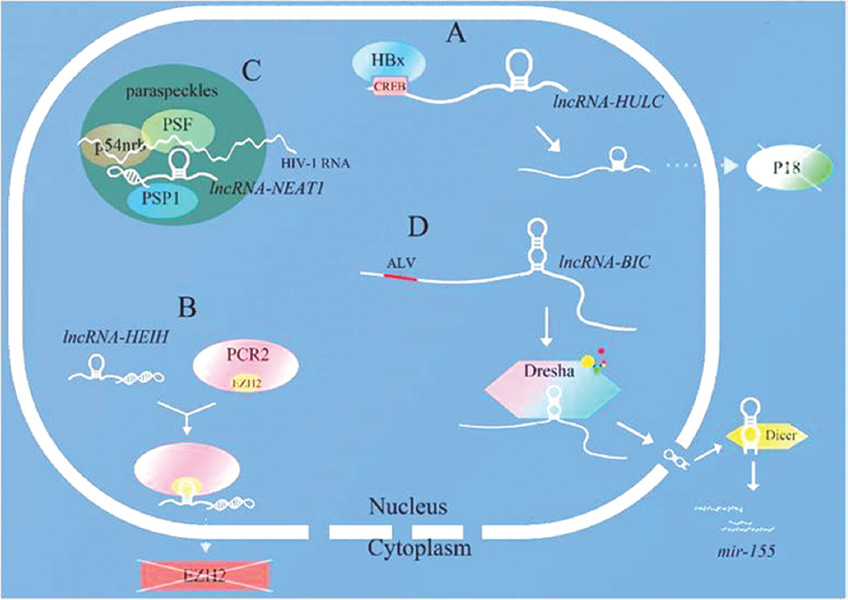


 - S01E13 Lift Me Up | Video clips by quotes | 82dfaa91 | 紗")





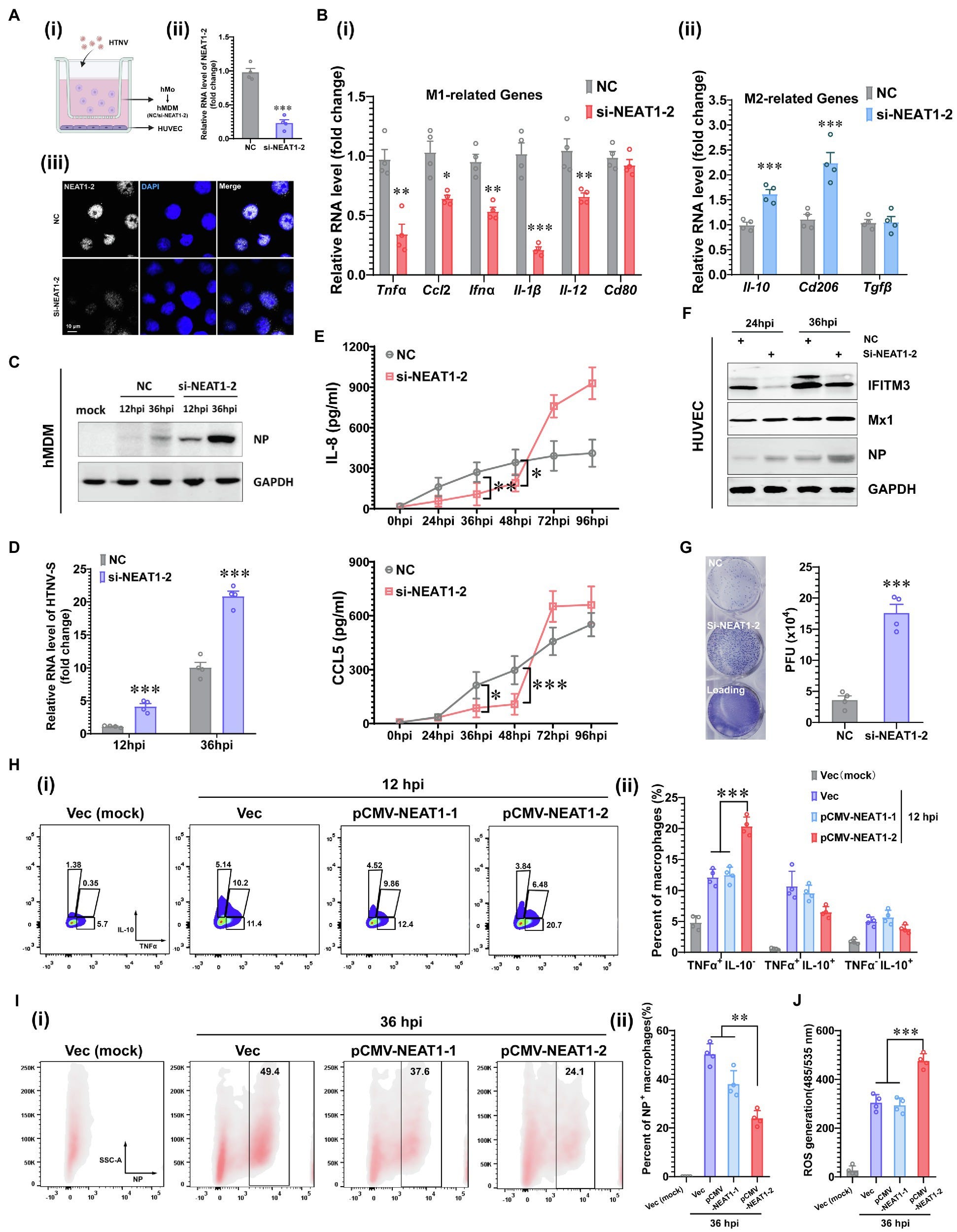


 Memedroid")





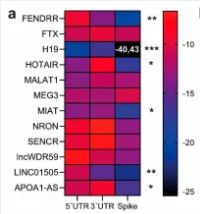












{kind=link}
{kind=link}
{kind=link}
Thanks for another research-laden post, Carp. I will send it to our "experts" and pollies.
Well, I’m 17% done, and bookmarked. This will have to be achieved in 3-5 sessions, considering my ‘Layman’ and ‘ESL (English was actually my fourth not second language, when I was already a in high school unfortunately)’ statuses.
Nonetheless, thank you, and very impressive so far.