


Spike protein (inc vax) induced immunodeficiency & carcinogenesis megathread #27: Further investigations into GP120, IL-6 & multidrug resistance

Any extracts used in the following article are for non commercial research and educational purposes only and may be subject to copyright from their respective owners.
Background

An already long & detailed Substack from January delved into the pathophysiology of brain fog, and the established literature signposted glycoprotein 120 as a major contributor:
GP120 inserts were confirmed a few weeks later by @Arkmedic when the unique sequences were found by using BLAST techniques and and also in a Moderna patent and several other papers:1

https://twitter.com/Jikkyleaks/status/1514185688610004993?s=19
Separately I reviewed IL-6 induced pathology, an oncogenic cytokine :
Quick summary: gp120 activates the p38 inflammatory MAPK pathway due to causing oxidative stress & potassium gating is interrupted leading to apoptosis.
This Substack explores the oncogenic potential of GP120, of how it upregulates cytokines including oncogenic interleukin 6, how both upregulate a multidrug resistant protein, the pathology of MRP1 and finally some potential therapeutics are explored.
Usual medical disclaimers apply: this is not medical advice, please seek guidance from a qualified medical professional before embarking on any treatment.
How the gp120 bearing S1 spike protein gets into the brain across the BBB to mediate brain fog is another question. Here's one route. Other non-viral routes may include via lipid nanoparticle trojan horses or exosomes. Data is hard to obtain but small amounts have been found in brain tissue of mice and cases of thrombosis are indicative:
The S1 protein of SARS-CoV-2 crosses the blood–brain barrier in mice (2020)
Abstract
It is unclear whether severe acute respiratory syndrome coronavirus 2, which causes coronavirus disease 2019, can enter the brain. Severe acute respiratory syndrome coronavirus 2 binds to cells via the S1 subunit of its spike protein. We show that intravenously injected radioiodinated S1 (I-S1) readily crossed the blood–brain barrier in male mice, was taken up by brain regions and entered the parenchymal brain space. I-S1 was also taken up by the lung, spleen, kidney and liver. Intranasally administered I-S1 also entered the brain, although at levels roughly ten times lower than after intravenous administration. APOE genotype and sex did not affect whole-brain I-S1 uptake but had variable effects on uptake by the olfactory bulb, liver, spleen and kidney. I-S1 uptake in the hippocampus and olfactory bulb was reduced by lipopolysaccharide-induced inflammation. Mechanistic studies indicated that I-S1 crosses the blood–brain barrier by adsorptive transcytosis and that murine angiotensin-converting enzyme 2 is involved in brain and lung uptake, but not in kidney, liver or spleen uptake.
https://www.nature.com/articles/s41593-020-00771-8
Thrombosis after covid-19 vaccination (2021)
“…However, in the absence of the results of study 514559, the biodistribution of ChaAdOx1 HBV in mice (study 0841MV38.001) confirms the delivery of vaccine into the brain tissues. The vaccine may therefore spur the brain cells to produce CoViD spike proteins that may lead to an immune response against brain cells, or it may spark a spike protein-induced thrombosis. This may explain the peculiar incidences of the fatal CVST observed with viral vector-based CoViD-19 vaccines. There is very little information in the public domain to assess the biodistribution of all genetic vaccines, however, it is anticipated that if it is characteristic to the viral vector employed in the vaccine, then the other vaccines using similar technology may also lead to the same safety concerns. Some examples of these vaccines include AstraZeneca/Oxford (Chimp adenoviral vector), J&J/Janssen (Human adenoviral vector 26), CanSinoBio (Human adenoviral vector 5), and Sputnik V (Human adenoviral vectors 26 and 5).
For COVID-19 mRNA Vaccine (Pfizer or Moderna), the biodistribution studies in animals were not conducted. The surrogate studies with luciferase and solid-lipid nanoparticles (Pfizer) confirm a biodistribution to the liver and other body tissues beyond the administration site [5]. For Moderna, the biodistribution of mRNA-1647 (encoding CMV genes) formulated in a similar lipid nanoparticulate delivery system confirms a biodistribution beyond the injection site, in particular, the distribution to the lymph nodes, spleen and the eye was noted [6]. However, the detailed tissue-specific distribution of mRNA vaccines encoding SARS-CoV-2 spike proteins (Pfizer or Moderna) is not fully known that can offer invaluable insights into the potential safety of these vaccines in peoples with pre-existing conditions or those on certain medications.”
Full article:
https://www.bmj.com/content/373/bmj.n958/rr-1
So we know that GP120 is in the spike protein sequence of both the virus and vaccines and that it can enter the brain in sufficient quantity to cause harm ie etiology of disease. Oxidative stress is indicated in the literature as the first step, but how exactly is this triggered? We have seen ROS generation before due to spike protein binding with ACE2, upregulating AngII and binding of Ang II to AT1R leads to increased ROS generation. This paper goes into some detail, but the pathway is quite different:
HIV-1 viral envelope glycoprotein gp120 produces oxidative stress and regulates the functional expression of multidrug resistance protein-1 (Mrp1) in glial cells
Patrick T. Ronaldson,Reina Bendayan
First published: 15 July 2008
Abstract
Brain human immunodeficiency virus type-1 (HIV-1) infection is associated with oxidative stress, which may lead to HIV-1 encephalitis, a chronic neurodegenerative condition. In vitro, oxidative stress can be induced in glial cells by exposure to HIV-1 envelope protein glycoprotein (gp120). Multidrug resistance proteins (Mrps) are known to efflux endogenous substrates (i.e. GSH and GSSG) involved in cellular defense against oxidative stress. Altered GSH/GSSG export may contribute to oxidative damage during HIV-1 encephalitis. At present, it is unknown if gp120 exposure can alter the functional expression of Mrp isoforms. Heat-shock protein 70, inducible nitric oxide synthase, intracellular GSSG, 2′,7′-dichlorofluorescein fluorescence, and extracellular nitrite were increased in primary cultures of rat astrocytes triggered with gp120, suggesting an oxidative stress response. RT-PCR and immunoblot analysis demonstrated increased Mrp1 mRNA (2.3-fold) and protein (2.2-fold), respectively, in gp120 treated astrocytes while Mrp4 mRNA or protein expression was not changed. Cellular retention of 2′,7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein, an established Mrp substrate, was reduced (twofold) in gp120-treated astrocytes, suggesting increased Mrp-mediated transport. In addition, GSH and GSSG export were enhanced in gp120-triggered cells. These data suggest that gp120 can up-regulate Mrp1, but not Mrp4, functional expression in cultured astrocytes. Our observation of increased GSH/GSSG efflux in response to gp120 treatment implies that Mrp isoforms may be involved in regulating the oxidative stress response in glial cells.
Abbreviations used
ABC
ATP-binding cassette
AM
acetoxymethyl ester
BCECF
2′, 7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein
BSA
bovine serum albumin
DCFH
2′,7′-dichlorofluorescein
DCFH-DA
2′,7′-dichlorofluorescein diacetate
HIV-1
human immunodeficiency virus type-1
HIVE
HIV-1 encephalitis
HSP70
heat-shock protein 70
IL
interleukin
iNOS
inducible nitric oxide synthase
JNK
c-Jun N-terminal kinase
MAPK
mitogen-activated protein kinase
MDR
multidrug resistance
MRP
multidrug resistance-associated protein
NO
nitric oxide
PBS
phosphate-buffered saline
P-gp
P-glycoprotein
ROS
reactive oxygen species
TNF
tumor necrosis factor
Astrocytes, the most abundant cell type in the CNS, perform several critical physiological functions, including blood–brain barrier homeostasis, regulation of axonal outgrowth, and formation of intracellular communication networks (Volterra and Meldolesi 2005). During pathological conditions, such as human immunodeficiency virus (HIV) encephalitis (HIVE), astrocytes are involved in immune signaling via synthesis and release of cytokines such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 (Persidsky and Gendelman 2003; Kaul et al. 2005; Speth et al. 2005). Cytokine secretion by astrocytes is believed to be triggered by soluble viral proteins [i.e. HIV-1 envelope glycoprotein (gp120)] that are released into the brain parenchyma (Kaul et al. 2005). Studies in cultured astrocytes suggest that direct binding of gp120 to chemokine receptors (i.e. CXCR4 and CCR5) at the astrocyte cell surface may mediate this inflammatory response (Wu et al. 1997; Koller et al. 2002). Recently, our laboratory has demonstrated CCR5-mediated production and secretion of TNF-α, IL-1β, and IL-6 in cultured astrocytes exposed to gp120 (Ronaldson and Bendayan 2006).
In addition to the inflammatory response, free radical production and oxidative stress are also involved in HIVE pathogenesis (Steiner et al. 2006; Reynolds et al. 2007). In particular, treatment of in vitro brain endothelial cell culture systems with gp120 has been reported to induce an oxidative stress response characterized by reduced intracellular concentrations of glutathione (GSH), reduced activity of the antioxidant enzymes GSH peroxidase and GSH reductase, and increased lipid peroxidation (Price et al. 2005, 2006). Studies in human mixed CNS cultures containing both glial cells and neurons have demonstrated that acute exposure to gp120 up-regulates inducible nitric oxide synthase (iNOS) expression with subsequent increases in nitric oxide (NO) production (Walsh et al. 2004). More recently, studies in cultured human astrocytes have shown that gp120 treatment increases the production of malondialdehyde, a lipid peroxidation byproduct and marker of oxidative stress (Visalli et al. 2007).
GSH is a critical factor responsible for the maintenance of cellular redox balance and antioxidant defense in the brain during oxidative stress. In the presence of reactive oxygen species (ROS), GSH is rapidly oxidized to glutathione disulfide (GSSG). Consequently, an elevated intracellular concentration of GSSG is indicative of oxidative stress. Astrocytes require high intracellular concentrations of GSH for cellular protection against a vast array of ROS (Minich et al. 2006). Previous studies have suggested that astrocytes may attempt to maintain redox balance during oxidative stress by the ATP-dependent extrusion of GSSG (Hirrlinger et al. 2001; Minich et al. 2006). Interestingly, these studies also reported that the same mechanisms involved in GSSG export may also be involved in the efflux of GSH (Hirrlinger et al. 2001; Minich et al. 2006). Membrane transport proteins that may be involved in cellular GSH/GSSG extrusion include the multidrug resistance (MDR) associated proteins (MRP in humans and Mrp in rodents), which are members of the ABCC subgroup of the ATP-binding cassette (ABC) superfamily of transporters. Several studies have demonstrated that both GSH and GSSG are substrates for multiple MRPs including MRP1/Mrp1 (Hirrlinger et al. 2001; Hirrlinger and Dringen 2005), MRP2/Mrp2 (Paulusma et al. 1999), and MRP4/Mrp4 (Rius et al. 2006). Although GSH transport has been reported to be elevated in Madin–Darby canine kidney cells transfected with MRP5 (Wijnholds et al. 2000), a recent study has shown that Mrp5 is not involved in the efflux of GSH or GSSG from primary cultures of rat astrocytes (Minich et al. 2006).
Several lines of evidence indicate that multiple MRPs are functionally expressed in glial cells. Functional isoforms for MRP1/Mrp1, MRP4/Mrp4, and MRP5/Mrp5 have been detected in rat and human astrocytes (Decleves et al. 2000; Hirrlinger et al. 2002a; Nies et al. 2004) and in rat microglia (Dallas et al. 2003, 2004). Although several studies have reported that negligible levels of MRP2/Mrp2 are expressed in the brain parenchyma (Choudhuri et al. 2003; Dallas et al. 2003), Hirrlinger et al. (2002a) detected the expression of Mrp2 mRNA in astrocytes isolated from embryonic rats. These discrepancies may be explained by differences in prenatal versus postnatal expression of Mrp2 (Kao et al. 2002; Yabuuchi et al. 2002).
Previous studies have reported that functional expression of MRP family members may be altered in response to oxidative stress. For example, induction of Mrp1 mRNA expression was observed in rat and human liver cell lines treated with pro-oxidants such as tert-butylhydroquinone, 2′,3′-dimethoxy-1′,4′-naphthoquinone, and menadione (Yamane et al. 1998). Mrp1 expression was also reported to be increased in heart (Jungsuwadee et al. 2006) and in brain (Joshi et al. 2005) isolated from C57BL/6 mice treated with adriamycin, a chemotherapeutic drug known to activate ROS production (Jungsuwadee et al. 2006). In primary cultures of rat astrocytes exposed to hydrogen peroxide, Hirrlinger et al. (2001) demonstrated an increase in Mrp1-mediated cellular efflux of GSH and GSSG, suggesting an upregulation of Mrp-mediated functional activity in response to oxidative stress (Hirrlinger et al. 2001). Interestingly, no change in Mrp1 gene and protein expression was observed in primary cultures of rat brain microvessel endothelial cells treated with hydrogen peroxide (Felix and Barrand 2002).
Recently, our laboratory has reported decreased functional expression of P-glycoprotein (P-gp) in cultured rat astrocytes treated with gp120 (Ronaldson and Bendayan 2006). However, it is unknown if other ABC transporters such as Mrp isoforms are altered in response to factors related to HIVE (i.e. gp120, oxidative stress). As we did not detect Mrp2 protein expression in our astrocyte cultures and Mrp3 and Mrp5 are not involved in the cellular extrusion of GSH/GSSG in cultured rat astrocytes (Homolya et al. 2003; Minich et al. 2006), we focused our work on Mrp1 and Mrp4. In particular, we have (i) evaluated oxidative stress markers and Mrp1, and Mrp4 functional expression in cultured rat astrocytes triggered with HIV-196ZM651 gp120 and (ii) investigated the effects of oxidative stress on the functional expression of these ABC transporters.
Discussion
ABC membrane transporters, such as P-gp and Mrp isoforms, are important determinants of xenobiotic and physiologic substrate permeation across the brain barriers (i.e. blood–brain barrier and blood-cerebrospinal fluid barrier) and brain cellular compartments (i.e. astrocytes, microglia). Until recently, the molecular expression (i.e. gene, protein) and functional activity of these transporters had primarily been characterized in non-pathological (i.e. healthy) cultures of rat astrocytes (Decleves et al. 2000; Ronaldson et al. 2004). To elucidate the role of brain pathologies associated with HIV-1 infection on the functional expression of ABC transporters, we implemented an in vitro astrocyte model of the inflammatory response associated with HIVE (Ronaldson and Bendayan 2006). In this model, cultured astrocytes were triggered with HIV-196ZM651 gp120 derived from a subtype C, R5-tropic HIV-1 viral strain. Using semiquantitative RT-PCR and ELISA analysis, we detected the increased production and secretion of cytokines (i.e. TNF-α, IL-1β, and IL-6) in primary cultures of rat astrocytes treated with HIV-196ZM651 gp120 (Ronaldson and Bendayan 2006). Brain concentrations of these cytokines are well-known to be significantly elevated during HIVE (Persidsky and Gendelman 2003; Kaul et al. 2005).
In addition to the innate immune response, oxidative stress may also play an important role in the pathogenesis of HIVE. In fact, decreased CNS concentrations of antioxidants such as GSH and increased levels of lipid peroxidation are documented in brain tissue from HIV-1 infected patients (Bains and Shaw 1997; Pocernich et al. 2000). The induction of oxidative stress and the production of ROS may be triggered by cellular exposure to HIV-1 viral proteins such as gp120. Several recent studies have shown that in vitro gp120 treatment may induce the expression of oxidative stress markers including increased iNOS protein levels and NO production (Walsh et al. 2004). Furthermore, decreased activity of the cellular antioxidant defense system has also been reported in response to in vitro gp120 exposure (Price et al. 2005). In this study, we examined the ability of gp120 to induce oxidative stress in primary cultures of rat astrocytes. When these astrocyte cultures were treated with HIV-196ZM651 gp120, we observed increased expression of HSP70. HSP70 is a member of the heat-shock family of proteins that are known to be up-regulated in response to cellular stress (Calabrese et al. 2004). In addition to this marker, we also reported the increased expression of iNOS in primary cultures of rat astrocytes triggered with HIV-196ZM651 gp120. The enzymatic conversion of l-arginine to l-citrulline is catalyzed by iNOS and yields NO as a byproduct of this reaction. NO can combine with superoxide anions to yield peroxynitrite, a potent cellular toxin. In vitro, NO has a half-life of only a few seconds but it rapidly reacts with oxygen and water to produce nitrites. Therefore, changes in NO production can be monitored by measuring nitrite concentrations in the extracellular milieu. In our hands, we observed increased nitrite concentrations in culture supernatants from astrocytes triggered with HIV-196ZM651 gp120 when compared with untreated controls. We also observed an increase in DCFH oxidation, which indicates an increase in cellular production of moderately reactive free radicals such as peroxides and superoxide. The cellular response to ROS involves GSH, a critical antioxidant that acts as a direct scavenger of ROS and as a cofactor in detoxification. During oxidative stress, GSH is rapidly oxidized to GSSG; consequently, a decrease in GSH and an increase in GSSG is indicative of an oxidative stress response (Price et al. 2005). In fact, the cellular GSSG : GSH ratio has been proposed to be an extremely sensitive indicator of oxidative stress (Neal et al. 1997). In our hands, we detected a significant decrease in intracellular GSH content and an increase in GSSG levels in response to HIV-196ZM651 gp120 treatment. Furthermore, we observed that the GSSG : GSH ratio was increased up to 194.1-fold after 24h treatment. Taken together, these data provide evidence for the induction of an oxidative stress response in cultured astrocytes triggered with HIV-196ZM651 gp120.
Recently, we observed significantly decreased P-gp protein expression in cultured astrocytes triggered with HIV-196ZM651 gp120 (4.7-fold) (Ronaldson and Bendayan 2006). Furthermore, the accumulation of two established P-gp substrates (i.e. digoxin and saquinavir) were significantly increased in HIV-196ZM651 gp120 cultures, suggesting a loss of P-gp-mediated transport activity (Ronaldson and Bendayan 2006). However, it was unclear if the molecular expression and/or functional activity of other ABC superfamily members (i.e. Mrp isoforms) were altered in response to gp120. As we did not detect Mrp2 protein expression in primary cultures of rat astrocytes and Mrp3/Mrp5 are not involved in GSH/GSSG efflux (Homolya et al. 2003; Minich et al. 2006), the present study focused on Mrp1 and Mrp4. We observed increased Mrp1 mRNA expression (2.3-fold) as well as increased Mrp1 protein expression (2.2-fold) in HIV-196ZM651 gp120 treated cultures. To our knowledge, this is the first report of altered Mrp expression in glial cells in response to treatment with an HIV-1 viral protein. In contrast, we did not observe any change in Mrp4 mRNA or Mrp4 protein in response to HIV-196ZM651 gp120 treatment.
…To determine if the observed changes in Mrp1 expression also corresponded to altered functional activity, we measured the cellular retention of BCECF, an established Mrp1, Mrp2, and Mrp4 substrate (Bachmeier et al. 2004; Wu et al. 2005). Our studies showed that HIV-196ZM651 gp120 treatment significantly reduced cellular BCECF retention at various time points in rat astrocyte monolayers suggesting an increase Mrp-mediated functional activity.
…Increased Mrp1 functional activity during pathological conditions such as HIVE may have dramatic physiological consequences for astrocytes, particularly via interference with antioxidant defense systems.
…In summary, this study describes the increased molecular expression and functional activity of Mrp1 in an in vitro model of an HIV-associated pathological condition. Our data provides evidence for the involvement of oxidative stress in the regulation of Mrp functional expression during a HIVE-associated pathophysiological response triggered by HIV-196ZM651 gp120. The enhanced efflux of GSH and GSSG, which may be related to increased functional expression of Mrp1, suggests that cellular redox status may change in response to HIV-196ZM651 gp120 exposure thus rendering glial cells more susceptible to oxidative cellular damage. Overall, these observations suggest that complex substrate-transporter interactions may occur during activation of oxidative stress in the brain and may contribute to the pathogenesis of CNS disorders such as HIVE.
Full paper:
https://onlinelibrary.wiley.com/doi/10.1111/j.1471-4159.2008.05479.x
I’m somewhat startled by some of the parallel findings in the paper below.
Let’s hope the rates for non-antiviral therapy CNS lymphoma and brain cancer are in no way representative:
ART=antiretroviral therapy
PLWH=People living with human immunodeficiency virus type 1
PCNSL=Primary Central Nervous System Lymphoma
“In pre-ART era, brain tumors were registered in 10% of PLWH [43]. Prevalence of PCNSL in AIDS patients was 3600-fold greater than in the general population, reaching 12% in AIDS patients.”
Its another long paper but I will highlight key passages.
By all means read it in full or come back to it later as new papers are published (or FOI’ed). You can even play Spike Protein Bingo and see how many matching pathways you can dab off. SNAIL is one (derived from zinc finger protein SNAI1), reverse transcription another, persistence in germinal centres another, and then we have lymphopenia & immune suppression through various pathologies. See my other posts from January for the rest.
SNAIL upregulation:
Spike protein (inc vax) induced immunodeficiency & carcinogenesis megathread #20: MiR-200, a new star miRNA in human cancer
This Substack relates several pathological processes, from phenotypical presentation to the TLR2-dependent activation of the NF-κB pathway, how SARS-CoV-2 spike protein induces transcriptional upregulation of Snail, how SNAIL opposes miR-200, how miR-200b & c work to suppress inflammation and act as tumor suppressors. MicroRNAs and carcinogenesis are also explored.
Hardly needing any introduction:
Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 mRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line
https://doorlesscarp953.substack.com/p/intracellular-reverse-transcription?utm_source=url&s=w
Oncogenic Effects of HIV-1 Proteins, Mechanisms Behind (2021)
Abstract
Simple Summary
People living with human immunodeficiency virus type 1 (HIV-1) (PLWH) are at increased risk of developing cancer despite successful antiretroviral therapy (ART). Here, authors suggest novel mechanism behind this phenomenon. HIV proteins, namely envelope protein gp120, accessory protein negative factor Nef, matrix protein p17, transactivator of transcription Tat and reverse transcriptase RT, are known to be oncogenic per se, to induce oxidative stress and to be released from the infected or expressing cells. These properties are proposed to underlie their capacity to affect bystander epithelial cells causing their malignant transformation, and to enhance tumorigenic potential of already transformed/cancer cells. HIV proteins can act alone or in collaboration with other known oncoproteins, specifically originating from the oncogenic human viruses such as human hepatitis B and C viruses, and human papilloma viruses of high carcinogenic risk, which cause the bulk of malignancies in people living with HIV-1 on ART.
Abstract
People living with human immunodeficiency virus (HIV-1) are at increased risk of developing cancer, such as Kaposi sarcoma (KS), non-Hodgkin lymphoma (NHL), cervical cancer, and other cancers associated with chronic viral infections. Traditionally, this is linked to HIV-1-induced immune suppression with depletion of CD4+ T-helper cells, exhaustion of lymphopoiesis and lymphocyte dysfunction. However, the long-term successful implementation of antiretroviral therapy (ART) with an early start did not preclude the oncological complications, implying that HIV-1 and its antigens are directly involved in carcinogenesis and may exert their effects on the background of restored immune system even when present at extremely low levels. Experimental data indicate that HIV-1 virions and single viral antigens can enter a wide variety of cells, including epithelial. This review is focused on the effects of five viral proteins: envelope protein gp120, accessory protein negative factor Nef, matrix protein p17, transactivator of transcription Tat and reverse transcriptase RT. Gp120, Nef, p17, Tat, and RT cause oxidative stress, can be released from HIV-1-infected cells and are oncogenic. All five are in a position to affect “innocent” bystander cells, specifically, to cause the propagation of (pre)existing malignant and malignant transformation of normal epithelial cells, giving grounds to the direct carcinogenic effects of HIV-1.
Keywords: human immunodeficiency virus type 1, epithelial cells, carcinogenicity, oxidative stress, reactive oxygen species, gp120, Tat, Nef, matrix protein p17, reverse transcriptase
2.1. Liver Cancer

The effect of HIV-1 on cells of the liver. Infection with HIV-1 and even exposition of hepatocytes (HP), hepatic stellate cells (HSC), Kupffer cells (KFC) to HIV-1 leads to production of reactive oxygen species (ROS) and induction of proinflammatory microenvironment, which in turn, promote/enhance replication of HBV, HCV, as well as HIV-1 itself, resulting in enhanced fibrosis, cirrhosis and development of hepatocellular carcinoma (HCC). Infections are depicted in red, secondary effects in dashed black, and events leading to tissue damage in ochre-colored lines.
2.2. Brain Cancer
PLWH are highly predisposed to developing brain cancer, including primary central nervous system lymphomas (PCNSL) and glioblastomas (GBM) [43,44]. In pre-ART era, brain tumors were registered in 10% of PLWH [43]. Prevalence of PCNSL in AIDS patients was 3600-fold greater than in the general population, reaching 12% in AIDS patients [44]. ART has dramatically reduced these rates, possibly due to the effect of protease inhibitors [45]. Still, the prevalence of brain tumors in PLWH appears to be higher than in general population: in USA; recorded prevalence of PCNSL in HIV-1 infected is 8.4% compared to <3.3% in the general US population [45,46] Also GBM occurs in PLWH (in various stages of HIV-1 infection) at a younger age and at a frequency 5.4- to 45-fold higher than in the general population [47]. Furthermore, the median survival rate in patients with GBM for PLWH is shorter than for HIV-1-negative patients receiving same treatment (an average of 8 compared to 14 months, respectively) [48].
The nature of the brain tumor-HIV-1 relationship is not fully understood. The majority of these tumors are central nervous system lymphomas but gliomas may develop as well. GBM tumors appear approximately three years after HIV-1 infection [43]. The stimulatory effect of HIV-1 infection on the development of GBM has been associated with reduced immune surveillance. However, survival of PLWH after GBM diagnosis is not associated with CD4+ cell counts [47]. The absence of a correlation between GBM development & progression with immune incompetence [47,49,50,51] indicates that aggressive tumor behavior is not a direct consequence of the immune deficiency and suggests direct involvement of HIV-1 in the initiation and progression of brain cancers. Importantly, HIV-1 infection in the brain is not limited to microglia/macrophages, but also affects astrocytes, which can then serve as a potential reservoir for further productive infection, viral persistence, and latency.
4. Potentiation of Carcinogenesis by Interactions of HIV-1 with Other Oncogenic Viruses
…One of the mechanisms of HIV-1 potentiation of liver cancer is the induction of oxidative stress. HCV, HIV-1 (and antiretroviral therapy) act together to activate production of ROS in HSCs and hepatocytes. ROS promote phosphorylation of the major mitogen-activated protein kinases active in human cells, p38 kinase, c-JUN N-terminal kinase (JNK) and extracellular signal-regulated kinase (ERK) that control cell growth, differentiation and apoptosis. In their turn, the phosphorylated p38 MAPK, JNK, and p42/44 ERK phosphorylate nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) protein complex, mastering transcriptional regulation of inflammation and cell death [31]. Following these events, phosphorylated NF-κB translocates to the nucleus, and where it normally modulates the production of both pro- and antifibrogenic/antiapoptotic genes, ensuring that liver cells are protected from apoptosis, but are capable to build the required inflammatory and immune responses [101]. In the presence of LPS, NF-κB can upregulate the expression of profibrogenic genes, such as procollagen α1, transforming growth factor β1 (TGF-β1) and tissue inhibitor of MMPs (TIMP-1) [31,101]. This process is accelerated by HIV-1/HIV-1 proteins: exposure of hepatocytes to HIV-1/HIV-1 proteins results in the elevated production of ROS and increased expression of collagen and TIMP1, further amplified by HCV infection, and even exposure to infectious HCV [35]. Taken together, these data indicate that HIV-1-mediated potentiation of hepatocellular carcinogenesis reflects a concerted action of HIV-1, HBV and HCV as viruses and/or individual viral proteins (Figure 1). Based on compelling data, McGivern & Lemon even suggested that the path to hepatocellular carcinoma in chronic hepatitis C shares important features with the carcinogenesis induced by HPV.
…Overall, these findings indicate that the increased incidence of AIDS-defining and non-AIDS defining forms of cancer in PLWH may reflect the direct or indirect, often concerted, carcinogenic effect(s) of HIV-1 and/or individual HIV-1 proteins on diverse infected as well as uninfected bystander cells. Furthermore, some HIV-1 proteins appear to be directly involved in cell transformation and propagation of malignant cells.
5. HIV-1 Antigens Involved in Cell Transformation and Tumor Propagation
5.2. Envelope Glycoprotein gp120
Glioma cells were shown to interact with the HIV-1 envelope protein gp120. This interaction promotes proliferation, migration, survival and stimulates glycolysis in glioma cell lines and tumor growth in animal models [127]. Increased glycolysis, also known as the Warburg effect characteristic of malignancy [128], results in increased protein and lipid synthesis, and promotes uncontrolled propagation (both proliferation and invasion) of tumor cells, as it provides them with glycolytic intermediary precursors required for the synthesis of DNA, proteins and lipids [127,129]. As Tat, gp120 induces EMT and cell migration through the TGF-B1 and MAPK signaling pathways.
5.4. Reverse Transcriptase (RT)
We have shown that constitutive expression of HIV-1 RT in murine mammary gland adenocarcinoma 4T1 cells leads to upregulation, in a concentration-dependent manner, of the expression of the transcription factors Twist and Snail tightly involved in EMT [134]. In vivo, expression of RT by 4T1 cells results in enhanced tumor growth and potentiates formation of metastasis in distal organs of immunocompetent syngenic mice [134]. Interestingly, this is not a common property of the reverse transcriptases, as constitutive expression of enzymatically active reverse transcriptase domain of telomerase reverse transcriptase, on contrary, suppressed both tumor growth and metastatic activity of 4T1 cells.
5.5. Matrix Protein p17
…Thus, several HIV-1 proteins are directly or indirectly oncogenic, stimulating transformation of healthy cells and propagation and aggressiveness of already existing cancer cells. These oncogenic properties are linked to two essential characteristics of these proteins: their capacity to induce oxidative stress with production of reactive oxygen species and their ability to exit HIV-1-infected cells (active or passive transport).
6. Oncogenic HIV-1 Proteins Induce Oxidative Stress
Virally-induced cancer evolves over long periods of time in the context of a strongly oxidative microenvironment, on the background of chronic inflammation. Oxidative stress induced by chronic viral infection is one of the factors driving neoplastic transformation, ultimately leading to oncogenic mutations in many cellular signaling cascades that drive cell growth and proliferation [42,141]. Oxidative damage of chromosomal DNA and chronic immune-mediated inflammation are key features of HBV, HCV, HPV, and HIV-1 infections [42,141]. As we have earlier reviewed, numerous lines of evidence show that HIV-1 infection triggers pronounced oxidative stress in both laboratory models and the context of in vivo infection by deregulation of oxidative stress pathways with escalation of ROS production and by inducing mitochondrial dysfunction [141]. As a result, PLWH exhibit multiple markers of oxidative stress including DNA damage [134,142]. The enhancement of ROS production is mediated by the envelope protein Gp120, Tat, Nef, RT, and p17.
6.2. Envelope Protein Gp120
Early findings indicated that gp120 increases free radical production from monocyte-derived macrophages (MDM) detected by spin-trapping methods, and that the spin trap adduct results from a reaction involving nitrogen oxide NO or its closely related oxidized derivatives [153]. We have earlier summarized a profound role of gp120 in the induction of oxidative stress [141], namely gp120 induces ROS production in cell lines of lymphoid origin, in the endothelial brain cells, astrocytes, neurons and microglia. In astrocytes, it enhances ROS production by several parallel mechanisms: via Fenton–Weiss–Haber reaction, NOX2 and NOX4, and cytochrome P450 2E1 (CYP2E1) [154,155]. The latter is mediated through the upregulation of CYP2E1 expression. In cancer (neuroblastoma) cells, gp120 induces proline oxidase that synthesized pyroline-5-carboxylate with concomitant generation of ROS (reviewed in [141]).
The effect of HIV-1/HIV-1 proteins on the cellular antioxidant defense system is controversial. They can both suppress and enhance antioxidant defense pathways [141]. Gp120 was shown to induce oxidative stress response. It up-regulates functional expression in cultured astrocytes of multidrug resistance protein 1 (Mrp1) which effluxes endogenous substrates glutathione and glutathione disulphide involved in cellular defense against oxidative stress [156]. It also upregulates the expression of nuclear factor erythroid derived 2-related factor 2 (Nrf2), a basic leucine zipper transcription factor which is known to regulate antioxidant defensive mechanisms) in human astrocytes, stimulating expression of key antioxidant defensive enzymes hemoxygenase (HO-1) and NAD(P)H dehydrogenase quinone1 (Nqo1) [157]. Pre-treatment of astrocytes with antioxidants or a specific calcium chelator BAPTA-AM, significantly blocks the upregulation of Nrf2, HO-1 and Nqo1.
6.4. Reverse Transcriptase
We have previously demonstrated that expression of RT by human cells induces production of ROS [163]. Later studies demonstrated that this is a property of different RT variants, including drug resistant variants, and variants retargeted for lysosomal processing and secretion [114,163]. Expression of all RT variants led to an increase in the levels of expression of Phase II detoxifying enzymes HO-1 and Nqo-1. Artificial secretion of RT resulted in a decrease of RT capacity to induce oxidative stress with a decrease in the production of ROS compared to the parental enzyme.
6.5. Matrix Protein p17
…Intrahippocampally injected into mice, p17 induced neurocognitive disorders, comparable in strenght to the effects of other known amyloidogenic proteins [166]. Interestingly, amyloidogenic proteins (typically amyloid-beta peptide Aβ) bound to redox active metal ions, such as copper, catalyse the production of ROS, in particular the most reactive one, hydroxyl radical. This effect may underlie the observed oxidative damage exerted by Aβ peptide on itself and on the surrounding molecules (proteins, lipids, DNA) [167]. One can hypothesize that matrix protein p17 with its amyloidogenic assemblies may trigger the production of ROS through a similar mechanism.
Thus, HIV-1 proteins with known oncogenic/mitogenic potential, Tat, gp120, Nef, RT, and potentially p17, have a potential to directly or indirectly induce oxidative stress, which could be one of the mechanisms by which they induce and potentiate carcinogenesis (Figure 3). Interestingly, HIV-1 proteins with an oncogenic potential involved in the induction of oxidative stress, such as Tat, gp120, Nef, RT, and possibly p17, can be found outside of the cells in which they are expressed.

Suggestive mechanism of direct carcinogenic effects of HIV-1 proteins. HIV-1 infected cells express and release gp120, Tat, Nef, p17, RT, each capable of the induction of oxidative stress. (1) p17 may trigger the production of ROS through binding of redox active metal ions by its amyloidogenic assemblies [167]. (2) Nef may indirectly activate NADPH oxidase by activating the Vav/Rac/p21-activated kinase (PAK) signaling pathway involved in phagocytic NADPH oxidase activation and produce peroxynitrite [160]. (3) Tat induces oxidative action through several independent mechanisms via NADPH oxidase, spermine oxidase (SMO) induction and mitochondrial dysfunction [148]. (4) RT induces ROS through unknown mechanisms. There is ROS –dependent activation of the Twist [134], which regulates the expression of Nrf2, which stimulating the expression of antioxidant enzymes (HO1, Nqol1). In addition, the Twist regulates the expression of the Snail. Both transcription factors, Twist and Snail, are involved in epithelial to mesenchymal transduction (EMT). (5) Gp120 increases free radical production from monocyte-derived macrophages (MDM) inducing nitrogen oxide (NO). In astrocytes (AS), it enhances ROS production by several parallel mechanisms: via cytochrome P450 2E1 (CYP2E1), NOX2 and NOX4, and the Fenton-Weiss-Haber reaction. Multidrug resistance proteins (Mrps) involved in cellular defense against oxidative stress. Mrp4 (isoform of Mrp) involved in the regulation of ROS and it acts against ROS [156]. In neuroblastoma cells (NB) gp120 was shown to induce proline oxidase that produces pyroline-5-carboxylate with a concomitant generation of ROS [141]. Production of ROS, which damage of bystander cells inducing oxidative damage of DNA, proteins and lipids, apoptosis and inflammation. DNA damage drives genomic instability and promotes transformation of healthy cells, and propagation and dissemination of malignant cells [168]. Arrows indicate: purple arrows—secretion/entering the intercellular space; black arrows—relationships and interactions; red arrows—production of ROS; blue arrows—oxidative stress response. Text above arrows designates the processes leading to the production of ROS, and text below the arrows, forms of ROS.
7. Oncogenic HIV-1 Proteins Inducing Oxidative Stress Are Found in the Extracellular Space
7.2. Envelope Protein gp120
Envelope protein gp120 is known to be secreted by chronically infected cells [180,181], particularly from the intraepithelial immune cells even in presence of ART [98]. A subset of PLWH demonstrate persistent circulation in plasma of gp120 [182] and in saliva [98]. Moreover, gp120 was found in tissues of PLWH [183]. Brain cells can be directly exposed to gp120 secreted by infiltrated and infected microglia and astrocytes [127]. Gp120 is internalized by bystander cells through receptor-independent mechanisms [184]. Internalization of gp120 leads to the release of several proinflammatory, angiogenic, and lymphangiogenic factors from affected cells.
7.4. Reverse Transcriptase (RT)
In our lab, we have shown secretion of RT into cell culture fluids of cells transiently expressing RT [114]. Recently, RT was also detected in the exosomes detected in the uterine of PLWH.
7.5. Matrix Protein p17
Thus, gp120 and Tat are actively secreted into the endothelial cell micro-environment, Nef can be neighboring uninfected cells including cells which cannot be infected with HIV-1, modulating their metabolism, cell cycle progression, ability to differentiate, motility, and, importantly, the genomic stability, through induction of ROS. Some HIV-1 proteins such as matrix p17 and gp120 can accumulate and persist in lymphoid tissues for at least 1 year after the on-start of ART on the background of successful suppression of viral replication [196]. These proteins are involved in different processes associated with malignant transformation and tumor growth with significant direct and indirect adverse effects on the epithelial cells. These include a range of responses that contribute to endothelial dysfunction, including enhanced adhesiveness, permeability, cell proliferation, apoptosis, as well as activation of cytokine secretion [86], eventually leading to malignant transformation (Figure 3). In this respect, their effect would resemble oncogenesis mediated by known viral oncoproteins originating from EBV, HTLV-1, KSHV, HCV, HBV, HPV, and identified as causative agents of both AIDS-defining and non-AIDS defining forms of cancer.
8. Conclusions
People living with human immunodeficiency virus receiving antiretroviral therapy are characterized by high prevalence of different forms of cancer affecting epithelial cells. HIV-1 does not infect epithelial cells, however both HIV virions and proteins were shown to be sequestered into epithelial cells and affect their functions. These proteins have three specific properties:
First, HIV proteins Tat, Nef, gp120, matrix protein p17, reverse transcriptase/RT induce oxidative stress with serious consequences in the form of DNA, protein and lipid damage, as well as changes in the intracellular signaling.
Second, Tat, Nef, gp120, matrix protein p17, RT have a direct carcinogenic potential as demonstrated in the series of in vitro experiments and experiments in the laboratory animals.
Third, Tat, Nef, gp120, matrix protein p17, reverse transcriptase/RT were shown to exit HIV expressing cells by different mechanisms, and, once present in the extracellular space, can be up-taken by innocent neighbor cells.
Sequestered/internalized by innocent bystander cells, these proteins modulate their metabolism, cell cycle progression, ability to differentiate, motility, redox balance (induce ROS) and genomic stability. Through this, they can trigger malignant transformation of normal cells. Another outcome is propagation (proliferation and dissemination) of already existing precancerous and cancer cells, and enhanced growth and metastatic activity of tumors expressing or exposed to HIV-1 proteins.
Altogether, we present a new mechanism of HIV-associated malignant transformation of epithelial cells driven by individual HIV proteins through the induction of reactive oxygen species. In this scenario, HIV-1 proteins act in a manner similar to the known viral oncogenes, and can cooperate with them promoting KSHV, EBV, HBV, HCV, and HPV-associated carcinogenesis. Such pathway of HIV associated carcinogenesis can co-occur together with carcinogenesis driven by persistent immune inflammation, and dysfunction of B cells, T cells and cellular components of the innate immune system.
Full paper:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7830613/
My conclusion. The last paragraph gives it away: “HIV-1 proteins act in a manner similar to the known viral oncogenes”.
Or to paraphrase, GP120 is persistent and pathogenic without requiring the virus itself, just as spike protein is persistent and pathogenic without requiring the virus itself. The twist being that the latter has the former as a biologically active inserts too.
The paper even classes GP120 as an “oncogenic HIV-1 protein”.
Read that again and remind yourself these biologically active inserts have been expressed in hundreds of millions of trusting transfected experimental gene therapy trial participants.
Further pathology
As the inserts are in the S1 spike protein, once cleaved at the FCS any free fragments don't need to bind to ACE2 to be endocytosed into a cell, which massively increases the type of tissue which can be affected. LNPs are also free of such restrictions of course. A recent study found that a percentage of monocytes were being depleted in a similar fashion due to exposure to spike protein.
GP120 is directly involved in CD4 depletion:
The CD4-gp120 interaction and AIDS pathogenesis
D J Capon et al. Annu Rev Immunol. 1991.
Abstract
Infection by the human immunodeficiency virus (HIV) leads to progressive destruction of the CD4+ subset of T lymphocytes, resulting in immunodeficiency and AIDS. The selectivity of CD4+ cell destruction is due to the specific binding of gp120, the external envelope glycoprotein of HIV, to CD4, initiating viral entry. Binding of gp120 to CD4 on the cell surface may also lead to CD4+ cell depletion by inappropriate immune targeting, and may interfere with CD4+ cell function and ontogeny by disrupting CD4-mediated cell signaling. The CD4-gp120 interaction is thus an obvious target for AIDS therapeutics.
https://pubmed.ncbi.nlm.nih.gov/1910691/
GP120 upregulates oncogenic IL-6 production:
HIV-1 gp120 Induces Expression of IL-6 through a Nuclear Factor-Kappa B-Dependent Mechanism: Suppression by gp120 Specific Small Interfering RNA (2011)
Abstract
In addition to its role in virus entry, HIV-1 gp120 has also been implicated in HIV-associated neurocognitive disorders. However, the mechanism(s) responsible for gp120-mediated neuroinflammation remain undefined. In view of increased levels of IL-6 in HIV-positive individuals with neurological manifestations, we sought to address whether gp120 is involved in IL-6 over-expression in astrocytes. Transfection of a human astrocyte cell line with a plasmid encoding gp120 resulted in increased expression of IL-6 at the levels of mRNA and protein by 51.3±2.1 and 11.6±2.2 fold respectively; this effect of gp120 on IL-6 expression was also demonstrated using primary human fetal astrocytes. A similar effect on IL-6 expression was observed when primary astrocytes were treated with gp120 protein derived from different strains of X4 and R5 tropic HIV-1. The induction of IL-6 could be abrogated by use of gp120-specific siRNA. Furthermore, this study showed that the NF-κB pathway is involved in gp120-mediated IL-6 over-expression, as IKK-2 and IKKβ inhibitors inhibited IL-6 expression by 56.5% and 60.8%, respectively. These results were also confirmed through the use of NF-κB specific siRNA. We also showed that gp120 could increase the phosphorylation of IκBα. Furthermore, gp120 transfection in the SVGA cells increased translocation of NF-κB from cytoplasm to nucleus. These results demonstrate that HIV-1 gp120-mediated over-expression of IL-6 in astrocytes is one mechanism responsible for neuroinflammation in HIV-infected individuals and this is mediated by the NF-κB pathway.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3119684/
You don't need GP120 to upregulate IL-6 but it helps. Spike protein induces upregulation for weeks or even months. There are several superantigenic motifs in addition to GP120 that can contribute to this.
On multidrug resistance
Not only does IL-6 promote tumor angiogenesis it also doubles the MRP1 multidrug resistance protein levels, just like we saw earlier with GP120 as an antioxidant defence. If you have an aggressive or other cancer that requires chemotherapy then this may be a problem for you.

“The drugs don't work” as the song goes:
Multidrug Resistance Proteins (MRPs/ABCCs) in Cancer Chemotherapy and Genetic Diseases (2011)
Abstract
The ATP-binding cassette (ABC) transporters are a superfamily of membrane proteins that are best known for their ability to transport a wide variety of exogenous and endogenous substances across membranes against a concentration gradient via ATP hydrolysis. There are seven subfamilies of human ABC transporters, one of the largest being the ‘C’ subfamily (gene symbol ABCC). Nine ABCC subfamily members, the so-called Multidrug Resistance Proteins (MRPs) 1-9, have been implicated in mediating multidrug resistance in tumor cells to varying degrees as the efflux extrude chemotherapeutic compounds (or their metabolites) from malignant cells. Some of the MRPs are also known to either influence drug disposition in normal tissues or modulate the elimination of drugs (or their metabolites) via hepatobiliary or renal excretory pathways. In addition, the cellular efflux of physiologically important organic anions such as leukotriene C4 and cAMP is mediated by one or more of the MRPs. Finally, mutations in several MRPs are associated with human genetic disorders. In this review article, the current biochemical and physiological knowledge of MRP1-MRP9 in cancer chemotherapy and human genetic disease is summarized. The mutations in MRP2/ABCC2 leading to conjugated hyperbilirubinemia (Dubin-Johnson syndrome) and in MRP6/ABCC6 leading to the connective tissue disorder Pseudoxanthoma elasticum are also discussed.
Keywords: ABCC Transporters, Dubin-Johnson Syndrome, Multidrug Resistance, Multiple Resistance Protein, Pseudoxanthoma Elasticum

…In tumor cells, the 190 kDa MRP1 can confer resistance to not only doxorubicin, but many other widely used antineoplastic drugs, including methotrexate (MTX), daunorubicin, vincristine and etoposide [5, 23, 29, 30]. Some of the newer so-called ‘targeted’ agents (e.g. certain tyrosine kinase inhibitors (TKIs)) that modify various signal transduction pathways, can also be transported by MRP1 [31]. MRP1 has been reported to be highly expressed in leukemias, esophageal carcinomas, and non-small cell lung cancer [32]. In addition, several reports have correlated MRP1 expression with clinical outcome [33]. Although, MRP1 mRNA and/or protein have been frequently detected in patient tumor samples, the overall contribution of this drug transporter to clinical drug resistance is still not well defined.
More:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3168698/
Influence of IL-6 on MDR and MRP-mediated multidrug resistance in human hepatoma cells
G Lee et al. Can J Physiol Pharmacol. 2001 Oct.
Abstract
The objective of this study was to examine effects of interleukin-6 (IL-6) on the expression and activity of the drug resistance transporters (MDR1 and MRP) in human hepatoma cell lines. Expression and activity of MDR1 and MRP transporters were examined in IL-6-treated and control HuH 7 and HepG2 cells using semi-quantitative RT-PCR analysis and by rhodamine 123 and 5-carboxyfluorescin efflux assays. Results from RT-PCR demonstrated expression of MRP3, MRP6, and MDR1 in HuH 7 cells and expression of MRP1, MRP2, MRP3, MRP6, and MDR1 in HepG2 cells. Compared with controls, treatment of HuH 7 cells with IL-6 (10 ng/mL, 24 h) resulted in a 1.8-fold increase in MRP-mediated efflux of 5-CF with a corresponding 1.5-fold induction of MRP3 mRNA levels (p < 0.05). Similarly, in HepG2 cells, a 2-fold increase in MRP functional activity and a 1.8-fold induction of MRP1 mRNA levels were seen in the IL-6 treated cells (p < 0.05). Treatment of cells with IL-6 was also found to cause significant reductions in the expression and activity of MDR1 in HuH 7 cells, but not in HepG2 cells. Our data suggest that IL-6 induces MRP expression and activity in human hepatoma cell lines. Suppressive effects of IL-6 on MDR1 expression and activity were also observed in HuH 7 cells. This underscores the importance of examining the regulation of multiple drug resistance proteins as these proteins may have opposing regulatory mechanisms in malignant cells.
https://pubmed.ncbi.nlm.nih.gov/11697747/
Regulation of Multidrug Resistance Protein 1 (Mrp1) by Tumor Necrosis Factor Alpha (TNF-α) in Cultured Glial Cells: Involvement of Nuclear Factor-κB (NF-κB) and c-Jun N-terminal Kinase (JNK) Signaling Pathways
Patrick T. Ronaldson, Tamima Ashraf and Reina Bendayan
Molecular Pharmacology January 5, 2010
Abstract
Pharmacotherapy of brain HIV-1 infection may be limited by ABC transporters [i.e., P-glycoprotein (P-gp), Multidrug Resistance Protein 1 (Mrp1)] that export antiretroviral drugs from HIV-1 brain cellular targets (i.e., astrocytes, microglia). Using an in vitro astrocyte model of an HIV-1 associated inflammatory response, our laboratory has shown that cytokines (i.e., TNF-α, IL-1β, IL-6), which are secreted in response to HIV-1 envelope glycoprotein gp120 exposure, can decrease P-gp functional expression; however, it is unknown if these same cytokines can alter expression and/or activity of other ABC transporters (i.e., Mrp1). In primary cultures of rat astrocytes, Mrp1 expression was increased by TNF-α (2.7-fold) but was not altered by IL-1β or IL-6. Cellular retention of BCECF, an Mrp substrate, was reduced in TNF-α treated astrocytes, suggesting increased Mrp-mediated transport. Pharmacological inhibition of NF-κB signaling with SN50 prevented both TNF-α release and Mrp1 expression changes in astrocytes triggered with gp120; however, SN50 did not attenuate Mrp1 expression in cells triggered with TNF-α. In contrast, Mrp1 functional expression was not altered in the presence of gp120 or TNF-α when astrocyte cultures were pre-treated with SP600125, an established JNK inhibitor. SP600125 did not affect TNF-α release from cultured astrocytes triggered with gp120. Mrp1 mRNA expression was increased after treatment with gp120 (1.6-fold) or TNF-α (1.7-fold), suggesting altered Mrp1 gene transcription. These data suggest that gp120 and TNF-α can up-regulate Mrp1 expression in cultured astrocytes. Furthermore, our results imply that both NF-κB and JNK signaling are involved in regulation of Mrp1 during an HIV-1 associated inflammatory response.
https://molpharm.aspetjournals.org/content/early/2010/01/05/mol.109.059410
The role of interleukin-6-STAT3 signalling in glioblastoma (2018)
Abstract
Glioblastoma is the most common type of malignant brain tumor among adults and is currently a non-curable disease due primarily to its highly invasive phenotype, and the lack of successful current therapies. Despite surgical resection and post-surgical treatment patients ultimately develop recurrence of the tumour. Several signalling molecules have been implicated in the development, progression and aggressiveness of glioblastoma. The present study reviewed the role of interleukin (IL)-6, a cytokine known to be important in activating several pro-oncogenic signaling pathways in glioblastoma. The current study particularly focused on the contribution of IL-6 in recurrent glioblastoma, with particular focus on glioblastoma stem cells and resistance to therapy.
Keywords: IL-6, STAT3, glioblastoma, therapeutics
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6144698/
Fibrosis is also a potential consequence of MRP1 upregulation, as touched on previously:
MicroRNA-9 limits hepatic fibrosis by suppressing the activation and proliferation of hepatic stellate cells by directly targeting MRP1/ABCC1 (2017)
Abstract
Liver fibrosis is a chronic liver disease characterized by the proliferation and activation of hepatic stellate cells (HSCs) and excessive deposition of extracellular matrix (ECM). Research suggests that microRNAs (miRNAs) are a new type of regulator of liver fibrosis. In the present study, we investigated the role of microRNA-9 (miR-9) in the process of liver fibrosis, as well as the underlying mechanism of action. Downregulated levels of miR-9 were found in fibrotic liver tissues and activated HSCs as detected by qRT-PCR; whereas, expression of multidrug resistance‑associated protein 1 (MRP1/ABCC1) was upregulated in the fibrotic liver tissues and activated HSCs. CCK-8 and BrdU assays revealed that miR-9 reduced the proliferative ability of the HSCs. In addition, expression levels of ECM-related genes (α-SMA, Col-1 and Timp-1), which are markers of HSC activation, were downregulated by miR-9. Conversely, an miR-9 inhibitor promoted cell proliferation and HSC activation. In addition, a luciferase reporter assay indicated that miR-9 targets the 3'-untranslated region (3'-UTR) of MRP1 and causes a significant decrease in MRP1. miR-9 inhibited the activation of the Hedgehog (Hh) pathway and the expression of MRP1, while this suppression was rescued by the overexpression of MRP1. Finally, a CCl4-induced mouse model of liver fibrosis was used to investigate the effects of miR-9 on liver fibrosis in vivo. The results showed that miR-9 abrogated hepatic fibrosis by suppressing the expression of MRP1 in CCl4-induced liver fibrotic mice. In conclusion, the present study demonstrated that miR-9 suppresses the proliferation and activation of HSCs through the Hh pathway by targeting MRP1, which suggests that miR-9 has therapeutic potential for liver fibrosis.
https://www.spandidos-publications.com/10.3892/or.2017.5382
Therapeutics
These are great results and not totally unexpected:
Hydroxychloroquine efficacy was detailed in the brain fog Substack and appears to have similar efficacy against MDR:
Hydroxychloroquine reverses the drug resistance of leukemic K562/ADM cells by inhibiting autophagy (2019)
https://pubmed.ncbi.nlm.nih.gov/31485616/
Ivermectin, a potential anticancer drug derived from an antiparasitic drug (2021)
Graphical abstract
Ivermectin has powerful antitumor effects, including the inhibition of proliferation, metastasis, and angiogenic activity, in a variety of cancer cells. This may be related to the regulation of multiple signaling pathways by ivermectin through PAK1 kinase. On the other hand, ivermectin promotes programmed cancer cell death, including apoptosis, autophagy and pyroptosis. Ivermectin induces apoptosis and autophagy is mutually regulated. Interestingly, ivermectin can also inhibit tumor stem cells and reverse multidrug resistance and exerts the optimal effect when used in combination with other chemotherapy drugs.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7505114/
CB2 and TRPV1 receptors mediate cannabinoid actions on MDR1 expression in multidrug resistant cells (2012)
Abstract
Background: Cannabis is the most widely used illicit drug in the world that is often used by cancer patients in combination with conventional anticancer drugs. Multidrug resistance (MDR) is a major obstacle in the treatment of cancer. An extensively characterized mechanism of MDR involves overexpression of P-glycoprotein (P-gp), which reduces the cellular accumulation of cytotoxic drugs in tumor cells.
Methods: Here we examined the role of cannabinoid receptors and transient receptor potential vanilloid type 1 (TRPV(1)) receptors in the effects of plant-derived cannabinoids on MDR1 mRNA expression in MDR CEM/VLB(100) cells which overexpress P-gp due to MDR1 gene amplification.
Results: We showed that both cannabidiol (CBD) and Δ(9)-tetrahydrocannabinol (Δ(9)-THC) (10 μM) transiently induced the MDR1 transcript in P-gp overexpressing cells at 4 but not 8 or 48 h incubation durations. CBD and THC also concomitantly increased P-gp activity as measured by reduced accumulation of the P-gp substrate Rhodamine 123 in these cells with a maximal inhibitory effect observed at 4 h that slowly diminished by 48 h. CEM/VLB(100) cell lines were shown to express CB(2) and TRPV(1) receptors. Δ(9)-THC effects on MDR1 expression were mediated by CB(2) receptors. The effects of CBD were not mediated by either CB(2) or TRPV(1) receptors alone, however, required activation of both these receptors to modulate MDR1 mRNA expression.
Conclusion: This is the first evidence that CB(2) and TRPV(1) receptors cooperate to modulate MDR1 expression.
https://pubmed.ncbi.nlm.nih.gov/22814029/
Cannabidiol (CBD) Inhibited Rhodamine-123 Efflux in Cultured Vascular Endothelial Cells and Astrocytes Under Hypoxic Conditions (2020)
Abstract
Despite the constant development of new antiepileptic drugs (AEDs), more than 30% of patients develop refractory epilepsy (RE) characterized by a multidrug-resistant (MDR) phenotype. The “transporters hypothesis” indicates that the mechanism of this MDR phenotype is the overexpression of ABC transporters such as P-glycoprotein (P-gp) in the neurovascular unit cells, limiting access of the AEDs to the brain. Recent clinical trials and basic studies have shown encouraging results for the use of cannabinoids in RE, although its mechanisms of action are still not fully understood. Here, we have employed astrocytes and vascular endothelial cell cultures subjected to hypoxia, to test the effect of cannabidiol (CBD) on the P-gp-dependent Rhodamine-123 (Rho-123) efflux. Results show that during hypoxia, intracellular Rho-123 accumulation after CBD treatment is similar to that induced by the P-gp inhibitor Tariquidar (Tq). Noteworthy, this inhibition is like that registered in non-hypoxia conditions. Additionally, docking studies predicted that CBD could behave as a P-gp substrate by the interaction with several residues in the α-helix of the P-gp transmembrane domain. Overall, these findings suggest a direct effect of CBD on the Rho-123 P-gp-dependent efflux activity, which might explain why the CBD add-on treatment regimen in RE patients results in a significant reduction in seizure frequency.
Keywords: cannabidiol, P-glycoprotein, hypoxia, endothelial cells, astrocytes
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7090129/
Glucosamine Reverses P-Glycoprotein-Mediated Multidrug Resistance in the Daunorubicin-Resistant Human Gastric Cancer Cells (2020)
Abstract
Glucosamine (GlcN) is a natural amino monosaccharide in the human body, and evidence of its anticancer effects is growing. In this study, we aimed to evaluate the effects of GlcN for its cytotoxicity, MDR reversal effects and inhibitory effects on function and expression of P-glycoprotein (P-gp) transporter in the daunorubicin-resistant human gastric cancer cells. Cell viability was measured by MTT assay to evaluate the cytotoxicity and multidrug resistance (MDR) reversal effects of GlcN. The effects of GlcN on function and mRNA expression level of P-gp transporter were assessed by flow cytometry and real-time RT-qPCR, respectively. Our results indicated that GlcN reduced the proliferation of human gastric cancer cell line EPG85-257 and its drug-resistant variant EPG85-257RD in a dose-dependent manner. GlcN (at the concentrations of 0.5 and 1 mM) also enhanced the sensitivity of EPG85-257RDB cells to daunorubicin. The cellular accumulation studies showed that GlcN inhibited efflux activity of P-gp and enhanced the mean fluorescent intensity of Rho123 while ˙it had no effects on P-gp gene expression in these cells. This study suggested that the inhibition of P-gp activity is a novel mechanism of action by which GlcN could reverse MDR in EPG85-257RDB cells.
https://pubmed.ncbi.nlm.nih.gov/31290701/
Metformin inhibits the development, and promotes the resensitization, of treatment-resistant breast cancer (2017)
Abstract
Multiple drug resistant (MDR) malignancy remains a predictable and often terminal event in cancer therapy, and affects individuals with many cancer types, regardless of the stage at which they were originally diagnosed or the interval from last treatment. Protein biomarkers of MDR are not globally used for clinical decision-making, but include the overexpression of drug-efflux pumps (ABC transporter family) such as MDR-1 and BCRP, as well as HIF1α, a stress responsive transcription factor found elevated within many MDR tumors. Here, we present the important in vitro discovery that the development of MDR (in breast cancer cells) can be prevented, and that established MDR could be resensitized to therapy, by adjunct treatment with metformin. Metformin is prescribed globally to improve insulin sensitivity, including in those individuals with Type 2 Diabetes Mellitus (DM2). We demonstrate the effectiveness of metformin in resensitizing MDR breast cancer cell lines to their original treatment, and provide evidence that metformin may function through a mechanism involving post-translational histone modifications via an indirect histone deacetylase inhibitor (HDACi) activity. We find that metformin, at low physiological concentrations, reduces the expression of multiple classic protein markers of MDR in vitro and in preliminary in vivo models. Our demonstration that metformin can prevent MDR development and resensitize MDR cells to chemotherapy in vitro, provides important medical relevance towards metformin’s potential clinical use against MDR cancers.
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0187191
Resveratrol-mediated reversal of tumor multi-drug resistance (2014)
Abstract
Multi-drug resistance (MDR) to cancer chemotherapy is a major obstacle to the effective treatment of tumors. Resveratrol, a natural product, may inhibit efflux transporters, such as P-glycoprotein (P-gp), multi-drug resistance-associated protein (MRP) and breast cancer resistance protein (BCRP), and could become a potential multi-drug-resistant regulator. But it remains unclear how resveratrol exerts its reversal effect. In this review, we attempt to reveal the interactions between resveratrol and ABC transporter proteins, and summarize the research profile of resveratrol's reversal mechanisms, thus to provide pivotal information on the development and application of multi-drug resistance reversal agents.
Curcumin down-regulates the multidrug-resistance mdr1b gene by inhibiting the PI3K/Akt/NF kappa B pathway (2008)
Abstract
Curcumin, a constituent of turmeric, has anti-inflammatory, anti-carcinogenic, and chemopreventive effects in several animal tumor models. The expression of P-glycoprotein (P-gp), encoded by the mdr gene, is often associated with multidrug resistance (MDR) to unrelated chemotherapeutic drugs in cancer cells. Here, we demonstrate that curcumin down-regulates P-gp expression in multidrug-resistant L1210/Adr cells. Transfection with a series of 5'-deleted constructs of the mdr1b gene promoter indicated that a proximal region between -205 and +42 of the sequence was responsible for the suppression of promoter activity by curcumin. This response might be associated with the inhibition of the phosphatidyinositol 3-kinase (PI3K)/Akt/nuclear factor-kappa B (NF-kappa B) signaling pathway by curcumin. Moreover, curcumin reversed the MDR of the L1210/Adr cells. Thus, curcumin can contribute to the reversal of the MDR phenotype, probably due to the suppression of P-gp expression via the inhibition of the PI3K/Akt/NF-kappa B signaling pathway.
Quercetin: a potential drug to reverse multidrug resistance (2010)
Abstract
This review centers on recent findings with respect to modulating cancer multidrug resistance (MDR) with the well-known flavonoid quercetin. After a short introduction of quercetin, major in vitro and in vivo findings are summarized showing that quercetin is a MDR modulator and thus a potential chemosensitizer. Finally, we contemplate future prospects of modulating MDR in the clinic.
…I will stop here, you can see the correlation!
BUT the exception here is NAC. This is not unexpected as it mediates the same antioxidant pathways as MRP via glutathione, very useful for countering spike protein induced ROS. However, it may not matter as NAC has anti-tumor properties of its own:
Cancer cell growth halted with cold and flu drug (2017)
“Researchers found that a medication used to ease symptoms of the common cold — called N-Acetylcysteine (NAC) — could also help to prevent the growth of cancer cells by depriving them of proteins that are important for their survival.”
https://www.medicalnewstoday.com/articles/320127
Pilot study demonstrating metabolic and anti-proliferative effects of in vivo anti-oxidant supplementation with N-Acetylcysteine in Breast Cancer (2017)
Abstract
Background
High oxidative stress as defined by hydroxyl and peroxyl activity is often found in the stroma of human breast cancers. Oxidative stress induces stromal catabolism, which promotes cancer aggressiveness. Stromal cells exposed to oxidative stress release catabolites such as lactate, which are up-taken by cancer cells to support mitochondrial oxidative phosphorylation. The transfer of catabolites between stromal and cancer cells leads to metabolic heterogeneity between these cells and increased cancer cell proliferation and reduced apoptosis in preclinical models. N-Acetylcysteine (NAC) is an antioxidant that reduces oxidative stress and reverses stromal catabolism and stromal-carcinoma cell metabolic heterogeneity, resulting in reduced proliferation and increased apoptosis of cancer cells in experimental models of breast cancer. The purpose of this clinical trial was to determine if NAC could reduce markers of stromal-cancer metabolic heterogeneity and markers of cancer cell aggressiveness in human breast cancer.
Methods
Subjects with newly diagnosed stage 0 and I breast cancer who were not going to receive neoadjuvant therapy prior to surgical resection were treated with NAC before definitive surgery to assess intra-tumoral metabolic markers. NAC was administered once a week intravenously at a dose of 150 mg/kg and 600 mg twice daily orally on the days not receiving intravenous NAC. Histochemistry for the stromal metabolic markers monocarboxylate transporter 4 (MCT4) and caveolin-1 (CAV1) and the Ki67 proliferation assay and TUNEL apoptosis assay in carcinoma cells were performed in pre- and post-NAC specimens.
Results
The range of days on NAC was 14–27 and the mean was 19 days. Post-treatment biopsies showed significant decrease in stromal MCT4 and reduced Ki67 in carcinoma cells. NAC did not significantly change stromal CAV1 and carcinoma TUNEL staining. NAC was well tolerated.
Conclusions
NAC as a single agent reduces MCT4 stromal expression, which is a marker of glycolysis in breast cancer with reduced carcinoma cell proliferation. This study suggests that modulating metabolism in the tumor microenvironment has the potential to impact breast cancer proliferation.
https://www.sciencedirect.com/science/article/abs/pii/S0093775417301367
N-acetylcysteine enhances multidrug resistance-associated protein 1 mediated doxorubicin resistance (2004)
Abstract
Background: Resistance of cancer cells against anticancer agents is caused partly by multidrug resistance-associated protein 1 (MRP1). The exact mechanism of MRP1-involved multidrug resistance has not yet been clarified, although glutathione (GSH) is likely to have a role for the resistance to occur. N-acetylcysteine (NAC) is a pro-glutathione drug. DL-buthionine (S,R)-sulfoximine (BSO) inhibits GSH synthesis. The aim of our study was to investigate the effect of NAC and BSO on MRP1-mediated doxorubicin resistance in human embryonic kidney (HEK293) and its MRP1-transfected 293MRP cells.
Materials and methods: Human embryonic kidney cells were transfected with a plasmid encoding the whole MRP1 gene. Both cells were incubated with doxorubicin in the presence or absence of NAC and/or BSO. The viability of both cells was determined under different incubation conditions. Glutathione, glutathione S-transferase (GST) and glutathione peroxidase (GPx) levels were measured in the cell extracts obtained from both cells incubated with different drugs.
Results: N-acetylcysteine increased the resistance of both cells against doxorubicin. DL-buthionine (S,R)-sulfoximine decreased NAC-enhanced MRP1-mediated doxorubicin resistance, indicating that induction of MRP1-mediated doxorubicin resistance depends on GSH synthesis. Doxorubicin decreased the cellular GSH concentration and increased GPx activity. Glutathione S-transferase activity was decreased by NAC.
Conclusion: Our results demonstrate that NAC enhances MRP1-mediated doxorubicin resistance and this effect depends on GSH synthesis. DL-buthionine (S,R)-sulfoximine seems a promising chemotherapy improving agent in MRP1 overexpressing tumour cells.
https://pubmed.ncbi.nlm.nih.gov/15473893/
Thank you for following.

There is a high likelihood the GP120 inserts are indeed biologically active: expressed, binding, superantigenic, oncogenic neurotoxins.
How this relates to ACE2 or TMPRSS2 binding is another question. I suspect spike protein binds to all 3 separately in various amounts depending on the cell type encountered.
The receptors are in place too:
CNS myelin induces regulatory functions of DC-SIGN–expressing, antigen-presenting cells via cognate interaction with MOG
"These data demonstrate that DC-SIGN is expressed on both microglia and perivascular macrophages in healthy human brain, closely associated with highly myelinated areas."
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4076586/https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4076586/
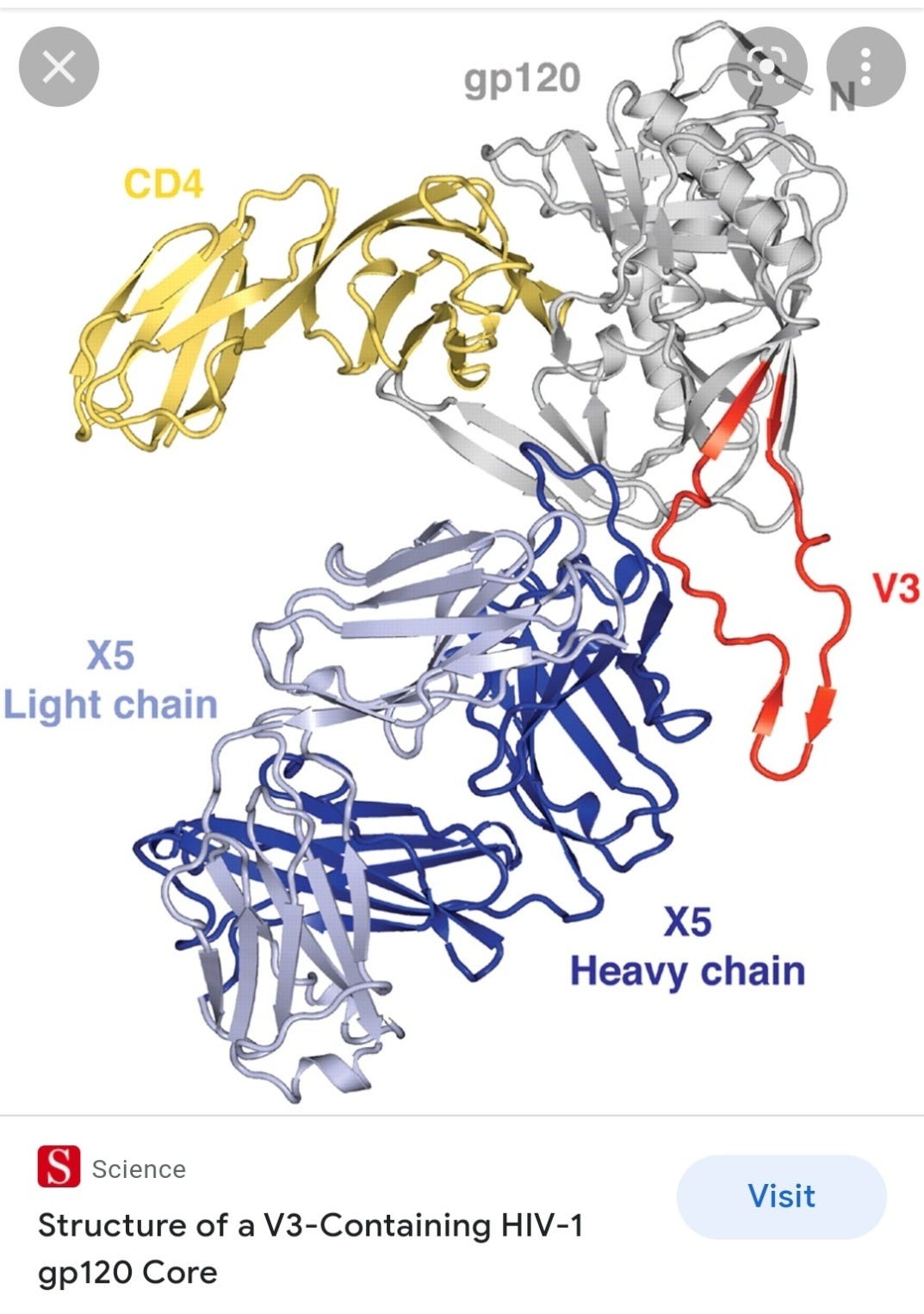
Structure of a V3-Containing HIV-1 gp120 Core (2005)
Abstract
The third variable region (V3) of the HIV-1 gp120 envelope glycoprotein is immunodominant and contains features essential for coreceptor binding. We determined the structure of V3 in the context of an HIV-1 gp120 core complexed to the CD4 receptor and to the X5 antibody at 3.5 angstrom resolution. Binding of gp120 to cell-surface CD4 would position V3 so that its coreceptor-binding tip protrudes 30 angstroms from the core toward the target cell membrane. The extended nature and antibody accessibility of V3 explain its immunodominance. Together, the results provide a structural rationale for the role of V3 in HIV entry and neutralization.
Wow.. It is a lot to take in. Very impressive work. Totally makes sense why the early meds & NAC, etc..were spoken against or pulled off shelves. Stop smoking campaign ads were constant. I am sickened that this was pushed on unsuspecting people, including most of my family. I wish they would read this article but deep down I know they won't. They recently got their 2nd booster after I begged all of them to wait for more info on these experimental toxins. Sadly my son & boyfriend got it to keep their jobs last summer. They have regretted it, and refuse to take anymore. They both got daily headaches, brain fog, breathing issues, & higher blood pressure after the 2nd shot. They have been on a weekly dose of ivermectin, & certain vitamins to feel normal again. The ivermectin helped almost immediately. No wonder the gov't agencies did a smear campaign on it & HCQ. Ivermectin not only can kill cancer, I believe I read in one of these articles it also kills HIV 1 plus many more things. I'm listening some articles that may be of interest
https://www.nature.com/articles/s41429-020-0336-z
https://www.nature.com/articles/ja201711
http://orthomolecular.org/resources/omns/v17n24.shtml
http://orthomolecular.org/resources/omns/v17n15.shtml