Any extracts used in the following article are for non commercial research and educational purposes only and may be subject to copyright from their respective owners.
This Substack also reviews some of the potential longer term risks of osteoporosis & musculoskeletal disorders in children, as these can be mediated by the same elevated inflammatory cytokines.
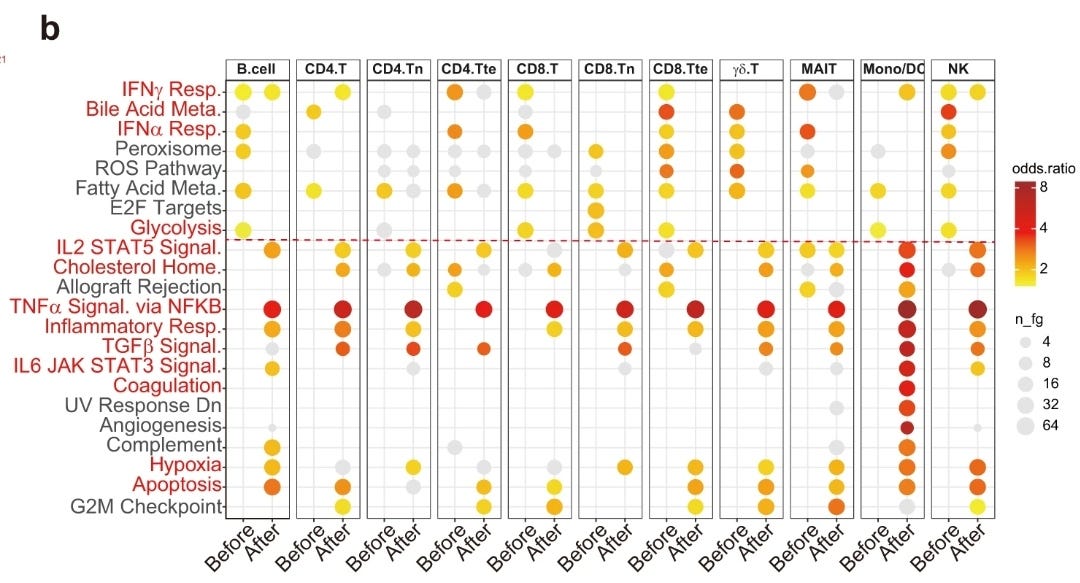
Note the extreme levels of NF-kB mediated TNFa elevation in the first paper and lL-1b in the second. Apart from the risks above, there are also potential oncogenic consequences, including a high risk of relapse for those in remission.
Comprehensive investigations revealed consistent pathophysiological alterations after vaccination with COVID-19 vaccines (2021)
Abstract
Large-scale COVID-19 vaccinations are currently underway in many countries in response to the COVID-19 pandemic. Here, we report, besides generation of neutralizing antibodies, consistent alterations in hemoglobin A1c, serum sodium and potassium levels, coagulation profiles, and renal functions in healthy volunteers after vaccination with an inactivated SARS-CoV-2 vaccine. Similar changes had also been reported in COVID-19 patients, suggesting that vaccination mimicked an infection. Single-cell mRNA sequencing (scRNA-seq) of peripheral blood mononuclear cells (PBMCs) before and 28 days after the first inoculation also revealed consistent alterations in gene expression of many different immune cell types. Reduction of CD8+ T cells and increase in classic monocyte contents were exemplary. Moreover, scRNA-seq revealed increased NF-κB signaling and reduced type I interferon responses, which were confirmed by biological assays and also had been reported to occur after SARS-CoV-2 infection with aggravating symptoms. Altogether, our study recommends additional caution when vaccinating people with pre-existing clinical conditions, including diabetes, electrolyte imbalances, renal dysfunction, and coagulation disorders.
Reduction of CD8+ T cells and increase in classic monocyte contents were exemplary. Moreover, scRNA-seq revealed increased NF-κB signaling and reduced type I interferon responses, which were confirmed by biological assays and also had been reported to occur after SARS-CoV-2 infection with aggravating symptoms. Altogether, our study recommends additional caution when vaccinating people with pre-existing clinical conditions, including diabetes, electrolyte imbalances, renal dysfunction, and coagulation disorders
Tumor necrosis factor and cancer, buddies or foes? (2009)
Abstract
Tumor necrosis factor (TNF) is a multifunctional cytokine that plays important roles in diverse cellular events such as cell survival, proliferation, differentiation, and death. As a pro-inflammatory cytokine, TNF is secreted by inflammatory cells, which may be involved in inflammation-associated carcinogenesis. TNF exerts its biological functions through activating distinct signaling pathways such as nuclear factor κB (NF-κB) and c-Jun N-terminal kinase (JNK). NF-κB is a major cell survival signal that is anti-apoptotic while sustained JNK activation contributes to cell death. The crosstalk between the NF-κB and JNK is involved in determining cellular outcomes in response to TNF. In regard to cancer, TNF is a double-dealer. On one hand, TNF could be an endogenous tumor promoter, because TNF stimulates cancer cells’ growth, proliferation, invasion and metastasis, and tumor angiogenesis. On the other hand, TNF could be a cancer killer. The property of TNF in inducing cancer cell death renders it a potential cancer therapeutic, although much work is needed to reduce its toxicity for systematic TNF administration. Recent studies have focused on sensitizing cancer cells to TNF-induced apoptosis through inhibiting survival signals such as NF-κB, by combined therapy. In this article we provide an overview of the roles of TNF-induced signaling pathways in cancer biology with specific emphasis on carcinogenesis and cancer therapy.
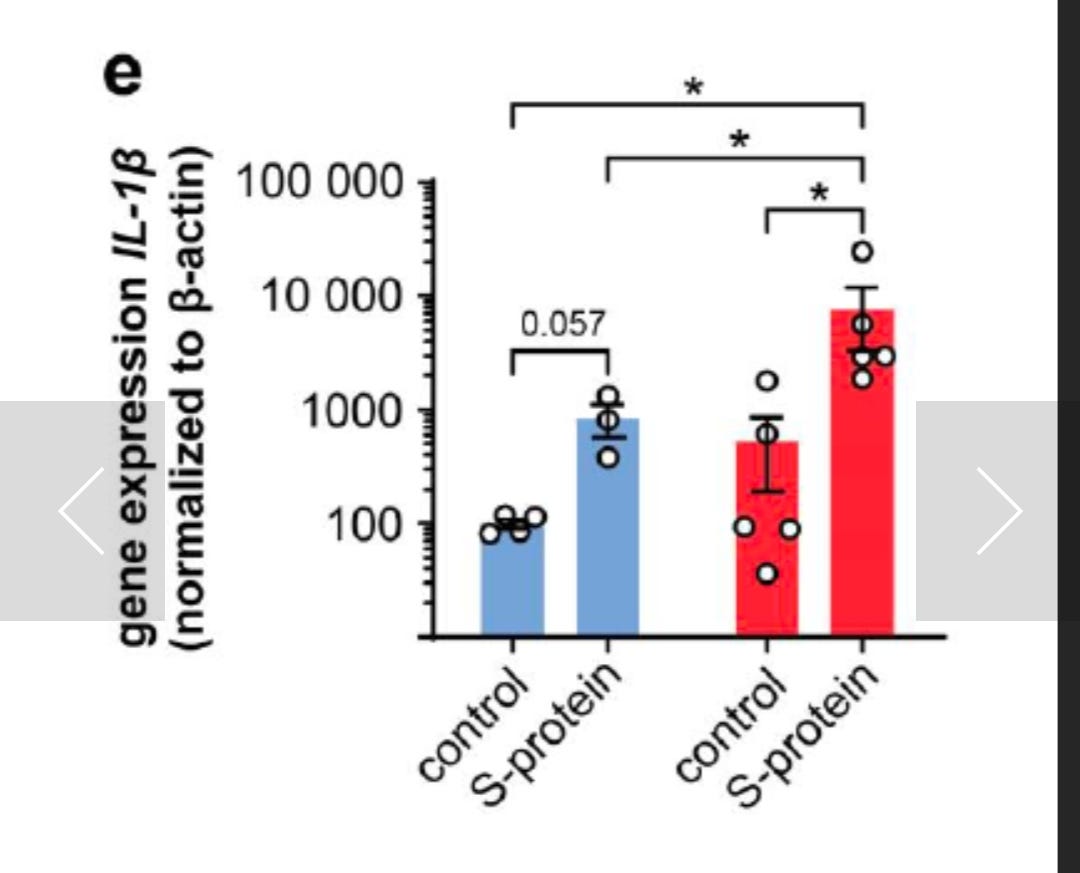
This preprint reports significant Interleukin-1-beta elevation.:
The SARS-CoV-2 spike protein primes inflammasome-mediated interleukin-1- beta secretion in COVID-19 patient-derived macrophages (2020)
Abstract
Innate immunity triggers responsible for viral control or hyperinflammation in COVID- 19 are largely unknown. Here we show that the SARS-CoV-2 spike protein primes inflammasome activation and interleukin 1-beta (IL-1β) secretion in macrophages derived from COVID-19 patients but not in macrophages from healthy SARS-CoV-2 naïve controls. Chemical NLRP3 inhibition blocks spike protein-induced IL-1β secretion ex vivo. These findings can accelerate research on COVID-19 vaccine design and drug treatment.
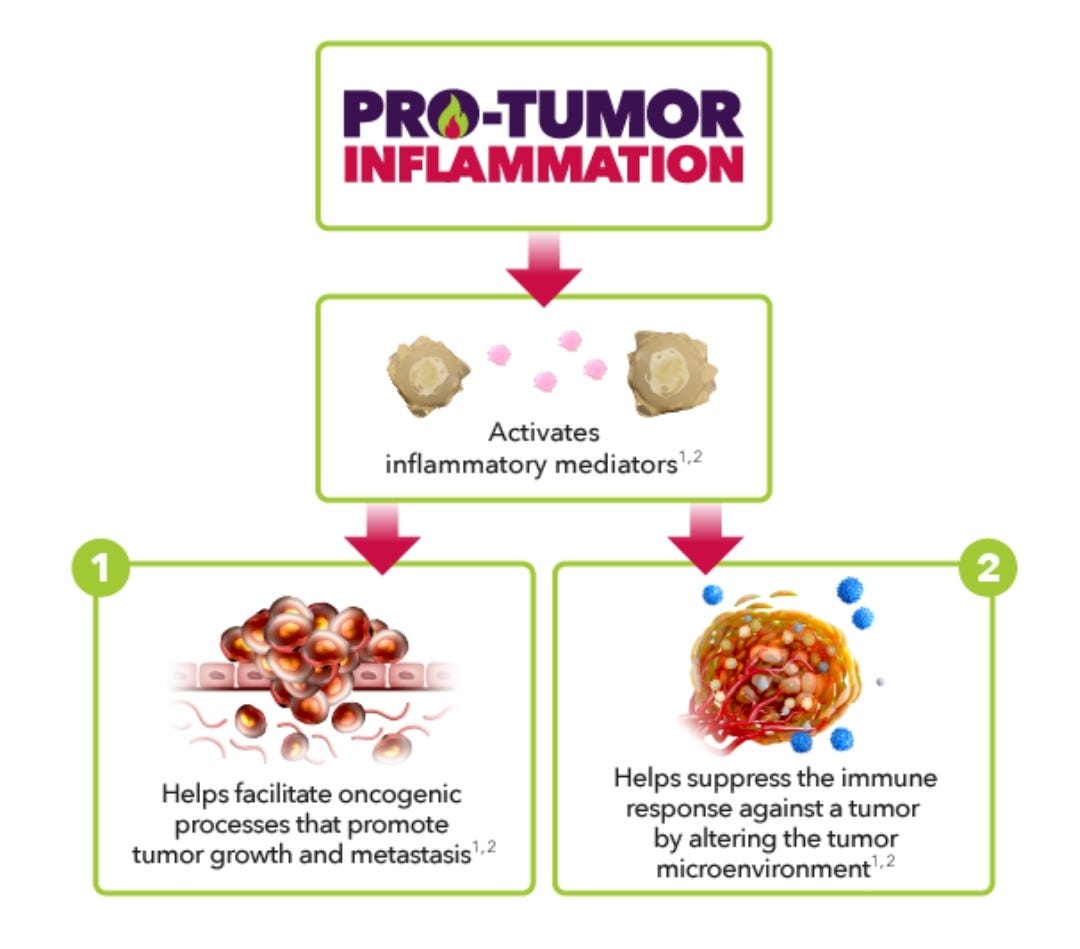
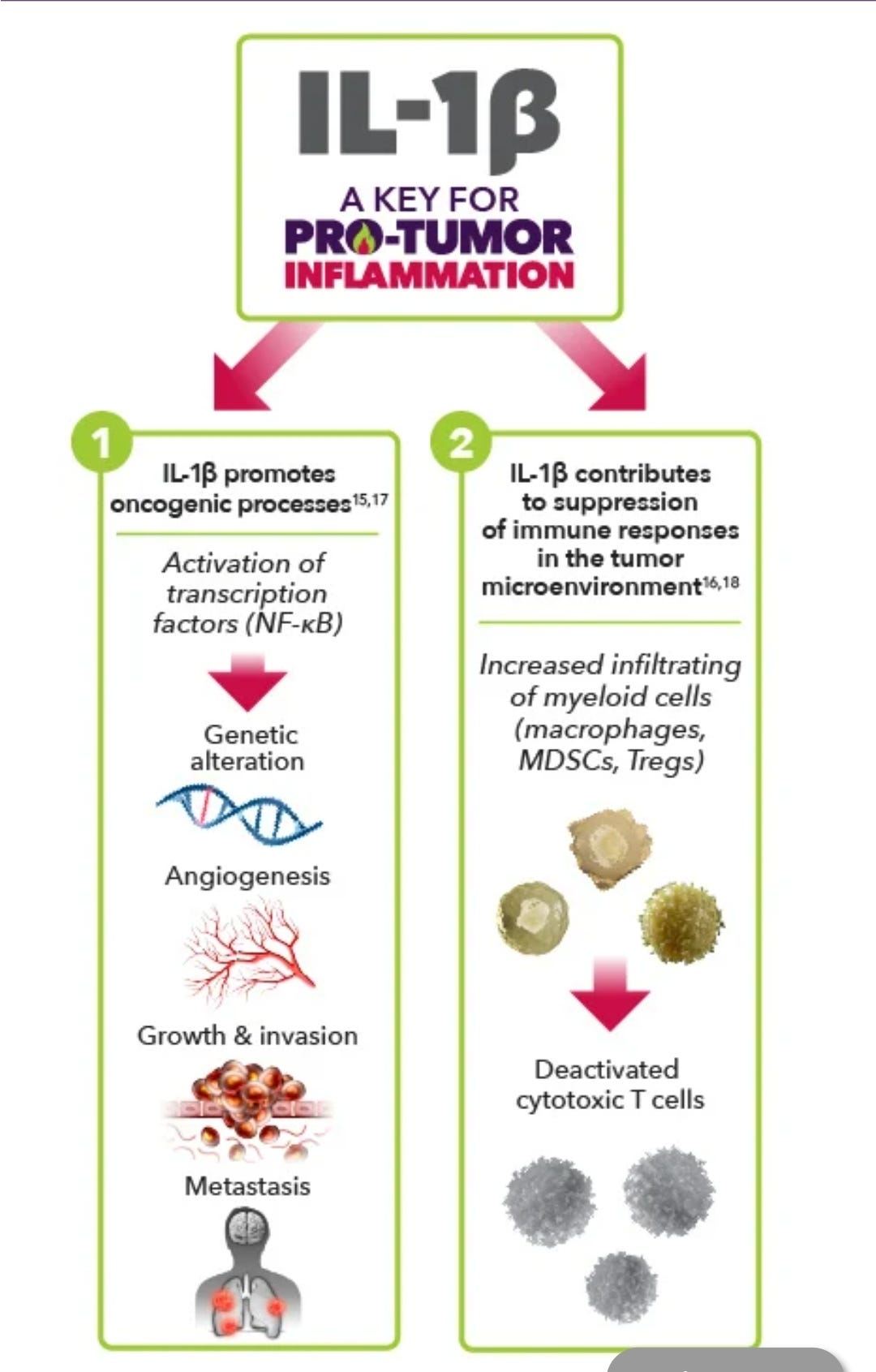
Pro-Tumor Inflammation refers to a type of unregulated inflammation that has many possible downstream consequences, such as facilitating tumor growth, survival, and metastasis
Osteoporosis is a health condition that weakens bones, making them fragile and more likely to break. It develops slowly over several years and is often only diagnosed when a fall or sudden impact causes a bone to break (fracture).
OSTEOBLASTS are the cells that form new bone. They also come from the bone marrow and are related to structural cells. They have only one nucleus. Osteoblasts work in teams to build bone. They produce new bone called "osteoid" which is made of bone collagen and other protein.
An osteoclast is a specialized cell that absorbs and removes bone, allowing for the development of new bone and maintenance of bone strength.
Hypoplasia: a condition of arrested development in which an organ or part remains below the normal size or in an immature state.
The growth plate, also known as the epiphyseal plate is a thin layer of cartilage that lies between the epiphyses and metaphyses, and is where the growth of long bones takes place.
CXCL10: Pro-inflammatory cytokine that is involved in a wide variety of processes such as chemotaxis, differentiation, and activation of peripheral immune cells, regulation of cell growth, apoptosis and modulation of angiostatic effects (PubMed:7540647, PubMed:11157474, PubMed:22652417).
Plays thereby an important role during viral infections by stimulating the activation and migration of immune cells to the infected sites (By similarity).
Mechanistically, binding of CXCL10 to the CXCR3 receptor activates G protein-mediated signaling and results in downstream activation of phospholipase C-dependent pathway, an increase in intracellular calcium production and actin reorganization (PubMed:12750173, PubMed:19151743).
In turn, recruitment of activated Th1 lymphocytes occurs at sites of inflammation (PubMed:12750173, PubMed:12663757).
Activation of the CXCL10/CXCR3 axis plays also an important role in neurons in response to brain injury for activating microglia, the resident macrophage population of the central nervous system, and directing them to the lesion site. This recruitment is an essential element for neuronal reorganization (By similarity).
Interleukin (IL): as discussed earlier, any of a group of naturally occurring proteins that mediate communication between cells. Interleukins regulate cell growth, differentiation, and motility. They are particularly important in stimulating immune responses, such as inflammation.
A recently published paper in the medical journal BONE associates significant levels of osteoporosis in mouse models with exposure to inflammatory cytokines, including TNF-a & interleukin 1-beta (IL-1β), both of which are also upregulated for a significant time period by transfection (up to 40 days in one study).
This was even found in mice showing no clinical signs of disease, indicating comparatively low cytokine levels were still associated with significant loss of bone density.
This potentially has profound implications for any vaccination of children, who are especially vulnerable as they are still undergoing musculoskeletal development.
Repeated annual or 6 monthly boosters will present the greatest risk of impediment to healthy development, as there is the very real risk of long term low level but significantly elevated pro-inflammatory cytokine & TNF levels. This may be compounded by LINE-1 reverse transcription or persistence in the lymph node germinal centres & monocytes, as has been demonstrated experimentally.
Apart from the elevated risk of fracture, potential growth retardation and deformities are described here in follow up studies of children who have undergone radio & chemotherapy:
A Systematic Review of Selected Musculoskeletal Late Effects in Survivors of Childhood Cancer (2015)
Osteoclast-mediated bone loss observed in a COVID-19 mouse model
(BONE, Volume 154, January 2022, 116227)
Highlights
•Mice surviving SARS-CoV-2 infection exhibit bone loss 12–14 days post-infection.
•Mice surviving SARS-CoV-2 infection exhibit increases in TRAP+ osteoclasts.
•Bone loss and elevated osteoclasts are seen in mice with no clinical signs of illness.
Abstract
The consequences of SARS-CoV-2 infection on the musculoskeletal system represent a dangerous knowledge gap. Aging patients are at added risk for SARS-CoV-2 infection; therefore, a greater understanding of the resulting musculoskeletal sequelae of SARS-CoV-2 infection may help guide clinical strategies. This study examined fundamental bone parameters among mice treated with escalating viral loads. Male C57BL/6J (WT, n = 17) and B6.Cg-Tg(K18-ACE2)2Prlmn/J mice (K18-hACE2 transgenic mice, n = 21) expressing human ACE2 (TG) were divided into eight groups (n = 4–6/group) and subjected to intranasal dosing of 0, 1 × 103, 1 × 104, and 1 × 105 PFU (plaque forming units) of human SARS-CoV-2. Animal health was assessed daily by veterinary staff using established and validated scoring criteria (activity, posture, body condition scores and body weight). We report here that mock and WT infected mice were healthy and completed the study, surviving until 12–14 days post infection (dpi). In contrast, the TG mice infected with 1 × 105 PFU all experienced severe health declines that necessitated early euthanasia (6–7 dpi). For TG mice infected with 1 × 104 PFU, 2 mice were also euthanized after 7 dpi, while 3 mice showed signs of moderate disease at day 6 dpi, but recovered fully by day 11 dpi. Four of the 5 TG mice that were infected with 1 × 103 PFU remained healthy throughout the study. This suggests that our study mimics what is seen during human disease, where some patients develop severe disease resulting in death, while others have moderate to severe disease but recover, and others are asymptomatic. At necropsy, femurs were extracted and analyzed by μCT. No difference was found in μCT determined bone parameters among the WT groups. There was, however, a significant 24.4% decrease in trabecular bone volume fraction (p = 0.0009), 19.0% decrease in trabecular number (p = 0.004), 6.2% decrease in trabecular thickness (p = 0.04), and a 9.8% increase in trabecular separation (p = 0.04) among surviving TG mice receiving any viral load compared to non-infected controls. No differences in cortical bone parameters were detected. TRAP staining revealed surviving infected mice had a significant 64% increase in osteoclast number, a 27% increase in osteoclast surface, and a 38% increase in osteoclasts per bone surface. While more studies are needed to investigate the long-term consequences of SARS-CoV-2 infection on skeletal health, this study demonstrates a significant reduction in several bone parameters and corresponding robust increases in osteoclast number observed within 2 weeks post-infection in surviving asymptomatic and moderately affected mice.
4. Discussion
Here we report that in a mouse model of COVID-19, infection of K18-hACE2 TG mice with SARS-CoV-2 represents 3 important classes of COVID patients: those who die (1 × 105 PFU), those with moderate to severe disease who may recover (1 × 104 PFU), and those who are asymptomatic (1 × 103 PFU). All of the infected TG mice experienced a robust loss of trabecular bone volume fraction (Fig. 4), likely owing to the significant 64% increase in TRAP+ osteoclasts (Fig. 6). Importantly, when we examined only mice that survived to the study endpoint (approximately 1 week longer survival duration), an even more dramatic change in trabecular bone parameters was observed. While additional studies would be required to test this idea, it is not an unreasonable hypothesis as it takes time for new osteoclasts to form and begin breaking down bone.
Another important observation is that the bone loss observed following SARS-CoV-2 infection occurs even in the surviving TG mice infected with 1 × 103 PFU, which had no clinical signs of illness as assessed by measuring changes in body weights, activity scores, posture scores, and BCS (Fig. 2, Fig. 3). The finding that all surviving mice in the 1 × 103 PFU group were as active as the controls, is important as if a mouse is sick and not moving, we may expect significant bone loss. Indeed, perhaps the most dramatic case scenario for inactivity and lack of mobility would be with unloading such as that observed with spaceflight or hindlimb unloading. In a hindlimb unloading study completed in the same background strain of mice at approximately the same age for the same study duration (4 month old C57BL/6 mice subjected to 2 weeks of unloading) investigators observed a 22% reduction in distal femur bone volume fraction [26]. In comparison, our surviving TG mice infected with 1 × 103 PFU of SARS-CoV-2 experienced a 25% reduction in distal femur bone volume fraction and remained load bearing throughout the study.
Although limitations of our study include low sample size (n = 4–6/genotype/viral dose), only one sex (male), and one age (approximately 19 weeks at the time of infection), as we are still in the midst of the COVID-19 pandemic, it was important to report on a statistically sound observation that has not been yet examined as a sequalae of this disease. This is especially true because as of September 16, 2021 the number of confirmed COVID-19+ cases reached over 226.9 million, while the death count was nearly 4.7 million [1]. Thus, there are millions of people who have already suffered from COVID-19 and bone loss may be yet another critically important secondary complication associated with having been infected with SARS-CoV-2.
Bone loss is a common side effect of aging and post-menopausal osteoporosis. Indeed, studies demonstrate that approximately 1 in 2 women, and 1 in 4 men over the age of 50 will suffer a fractured bone as a result of osteoporosis [27]. Therefore, it is formally possible that those who contracted COVID-19 may be more likely to be diagnosed with osteoporosis and/or may be at higher risk of developing fractures. However, it is important to note that these studies do not demonstrate whether the bone loss observed can be recovered over time. While it would be important to determine this for all individuals, it may be particularly important to understand this for the aged population, where they are already at risk for age-associated osteoporosis.
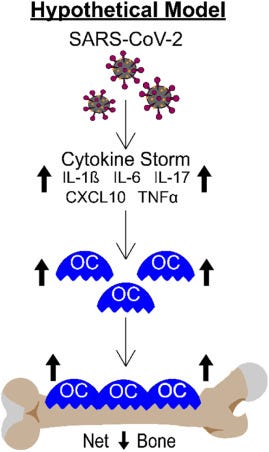
It is critically important to understand the long-term consequences of this dramatic bone loss. However, it is equally important to understand the mechanisms of action. While not specifically tested in this study, it is well known that patients with severe forms of COVID-19 have upregulated expression of numerous cytokines and growth factors which together lead to the inflammatory “cytokine storm” [[28], [29], [30], [31], [32], [33]]. Many of these cytokines, including IL-1β, IL-6, IL-17, CXCL10, and TNF-α are known to regulate osteoclastogenesis and/or bone resorption [[3], [4], [5], [6], [7], [8]], suggesting these factors may contribute to the SARS-CoV-2 stimulated bone loss observed in our K18-hACE2 mouse model. Fig. 7 shows a working model of how SARS-CoV-2 infection may be regulating osteoclast-mediated bone loss.
Fig. 7. Schematic demonstrating our hypothetical model describing how the cytokine storm associated with SARS-CoV-2 infection may result in osteoclast-mediated bone loss.
We recognize that the cytokine storm is just one possible mechanism by which bone loss occurs and much more work is needed to fully elucidate the mechanisms of action. For example, a recent study found that SARS-CoV-2 could directly infect bone marrow macrophages to a higher extent than mature osteoclasts [34]. Further, their study suggests that infection may alter the ability of bone marrow macrophages to differentiate into osteoclasts. Regardless of the mechanism of action, infection with SARS-CoV-2 results in a dramatic upregulation in osteoclastogenesis and a robust loss of bone within 2 weeks post-infection in a COVID-19 mouse model. Whether these findings translate into humans remains to be determined, but if they do, an increase in fractures may be observed secondary to COVID-19 disease in the not too distant future.
SARS-CoV-2 can alter bone marrow macrophage (BMM)-to-osteoclast differentiation, study finds (2021)
The coronavirus disease 2019 (COVID-19) pandemic, caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), is primarily a respiratory illness.
However, as the pandemic evolves, scientists have observed a range of different ways that the virus can affect the body. Alongside the lungs, SARS-CoV-2 has been seen to affect other organs, including the heart, brain and gastrointestinal tract.
Now, a team of researchers in China and Australia has shown that SARS-CoV-2 infection can affect bone marrow macrophage (BMM)-to-osteoclast differentiation, which may impact the skeletal system.
Study: Neuropilin-1 Mediates SARS-CoV-2 Infection in Bone Marrow-derived Macrophages. Image Credit: sciencepics / Shutterstock
In the current study, which appeared on the pre-print server bioRxiv*, the team found that authentic SARS-CoV-2 could efficiently infect both human and mouse BMMs and alter the expression of macrophage chemotaxis and osteoclast-related genes.




Sheeeeee-it! Is everyone going to crumble before they die?