A Computational Study of Ivermectin and Doxycycline Combination Drug Against SARS-CoV-2 Infection;
What’s stopping doctors repurposing generic medicines? Answer - not a lot!
23rd December ‘23: Caution added for doxycycline.
Any extracts used in the following article are for non commercial research and educational purposes only and may be subject to copyright from their respective owners.
Substack limitations
Once Substacks’ support the use of internal hyperlinks I will add them to the abstract. Otherwise please use a “Find In Page” keyword search to navigate or click on the footnote text numbers.
If received via email I recommend clicking on the hyperlinked title to read the latest correctly formatted version in a browser. Unfortunately it is not possible to email out revised versions.
Abstract
This review begins with a brief introduction to doxycycline as a drug: its uses, side effects, history and chemical formula with the 5-ringed structure typical of the tetracyclines.
Doxycycline and its iron chelation mechanisms and effects are then described.
In 2021, Faure et al performed in vitro experiments to investigate the synergistic interactions between five tetracyclines and tobramycin with an iron chelator (CP762) against two reference strains and nine clinical isolates of Pseudomonas aeruginosa from cystic fibrosis patients.
They found that as it binds with high affinity to iron this inhibited its antibacterial effects by competing with the magnesium binding site on the bacterial ribosome. The addition of another iron chelator, CP762, synergistically restored the magnesium bridge binding.
In 1999 Alkawash et al appeared to find lactoferrin/doxycycline antibacterial synergy, and by a large margin of 32 to 64 fold against B. cepacia.
In 2015, Wu et al investigated its effects in vitro on the replication of vesicular stomatitis virus.
Of particular note here for also treating long covid/vaccine sequalae is that doxycycline acts as both an antiviral and an anticancer therapeutic agent by the induction of expression of the key tumor suppressor p53.
Induction of p53 expression by doxycycline is then discussed in more detail. Son et al (2009) conducted an in vitro investigation and concluded that the tumor suppressive effects on pancreatic cancer cells were due to the activation of proapoptotic genes, inhibiting IL-8 expression, and suppression of antiapoptotic genes.
In 2020, Mosquera-Sulbaran and Hernández-Fonseca published a review on the use of tetracycline as an anti COVID-19 therapeutic.
2 clinical trials using doxycycline and ivermectin are then discussed. No results were available from the first of these, but from the second the only participants to die of COVID-19 were 3 from the placebo group of 200. Duration and severity of symptoms in the treatment group were also significantly reduced.
Three papers investigating iron chelation, inhibition of tumors and metastasis are reviewed. Buss et al (2003) recognized the potential of using iron chelation in cancer therapy and their possible synergistic effects.
From 2013, Richardson et al review how the iron chelator DFO can inhibit key signalling pathways which induce epithelial mesenchymal transition (EMT) in pancreatic cancer and other tumors. EMT is described.
A paper by Morales and Xue (2021) reviews the targeting of iron metabolism in cancer therapy.
Ubiquitination is described, as is evidence for the HIF inhibitory effects of both lactoferrin and ivermectin. This is important for working synergistically with doxycycline to help avoid resistance from cancer cells.
To complete this review, four papers discuss the anticancer properties of doxycycline.
In 1998, Fife et al found that, in vitro, doxycycline can significantly inhibit the growth of prostate and breast cancer tumors by the inhibition of matrix metalloproteinases (MMPs) and induction of apoptosis.
From 2016, Zhang et al conducted an in vitro study using human breast cancer cell lines.
A paper by Zhu et al (2017) conducted an in vitro investigation into how doxycycline synergizes with the chemotherapeutic drug doxorubicin to inhibit the proliferation of castration-resistant prostate cancer cells, a condition that was previously untreatable.
And from 2019, Markowska et al conducted a review into the repositioning of doxycycline, salinomycin, monensin and ivermectin as cancer drugs.
To conclude this Substack, dosing and contraindications for doxycycline monohydrate are considered.
Appendix:
A Computational Study of Ivermectin and Doxycycline Combination Drug Against SARS-CoV-2 Infection;
What’s stopping doctors repurposing generic medicines? Answer - not a lot
What is Doxycycline?
(6-deoxy-hydroxytetracycline)
Common brand names: Alodox, Doryx, Mondoxyne
Doxycycline is a broad-spectrum tetracycline-class antibiotic used in the treatment of infections caused by bacteria and certain parasites. It is used to treat bacterial pneumonia, acne, chlamydia infections, Lyme disease, cholera, typhus, and syphilis. It is also used to prevent malaria in combination with quinine. Doxycycline may be taken by mouth or by injection into a vein.
Common side effects include diarrhea, nausea, vomiting, abdominal pain, and an increased risk of sunburn. Use during pregnancy is not recommended. Doxycycline is a broad-spectrum antibiotic, of the tetracycline class. Like other agents of the tetracycline class, it either slows or kills bacteria by inhibiting protein production. It kills malaria by targeting a plastid organelle, the apicoplast.
Doxycycline was patented in 1957 and came into commercial use in 1967. It is on the World Health Organization's List of Essential Medicines. Doxycycline is available as a generic medicine. In 2019, it was the 90th most commonly prescribed medication in the United States, with more than 8 million prescriptions
Chelation is a type of bonding of ions and molecules to metal ions. It involves the formation or presence of two or more separate coordinate bonds between a polydentate (multiple bonded) ligand and a single central metal atom. These ligands are called chelants, chelators, chelating agents, or sequestering agents. They are usually organic compounds, but this is not a necessity, as in the case of zinc and its use as a maintenance therapy to prevent the absorption of copper in people with Wilson's disease.
Chelation is useful in applications such as providing nutritional supplements, in chelation therapy to remove toxic metals from the body, as contrast agents in MRI scanning, in manufacturing using homogeneous catalysts, in chemical water treatment to assist in the removal of metals, and in fertilizers
In 2021, Faure et al performed in vitro experiments to investigate the synergistic interactions between five tetracyclines and tobramycin with an iron chelator (CP762) against two reference strains and nine clinical isolates of Pseudomonas aeruginosa from cystic fibrosis patients.
They found that as tetracyclines bind with high affinity to iron this inhibits their antibacterial effects by competing with the magnesium binding site on the bacterial ribosome. The addition of another iron chelator, CP762, synergistically restored the magnesium bridge binding.1
Lactoferrin is also an iron chelator, so it may be postulated that a similar synergism exists as with CP762.
Highlights
• High affinity iron chelation enhances the antibacterial activity of tetracyclines.
• High affinity iron chelation synergises with doxycycline against P. aeruginosa.
• Doxycycline chelates iron and loses antibacterial activity.
• Iron chelation re-establishes the susceptibility of iron bound doxycycline.
• Iron chelation enhances doxycycline activity in a biofilm setting.
Results
There was synergy between CP762 and all tetracyclines, except minocycline, against the reference strains but that against clinical isolates was variable. Synergy was not demonstrated for tobramycin against any of the strains tested. This led to the hypothesis that iron chelation preserves the binding of tetracyclines to the bacterial ribosome. Susceptibility to iron-bound doxycycline was decreased by two- to four-fold and synergistic interactions with the iron chelator were consistently more intense with iron-bound doxycycline than with doxycycline alone. The doxycycline–iron chelator combination also significantly reduced cell viability in established biofilms.
Tetracyclines are established chelators, with a high affinity for copper, iron and zinc, and lower affinities for magnesium and calcium. Tetracyclines bind to the 30S bacterial ribosome through a magnesium bridge. Abundant metals, such as iron, may interfere with this mechanism by binding to the magnesium binding site. The working hypothesis for the current study is that CP762 sequesters iron thereby minimising iron-binding to other ligands, e.g., tetracycline. This will promote complexation with lower affinity ions, such as magnesium, required for binding to the bacterial ribosome.
In conclusion, iron chelation through CP762 was found to enhance the activity of five antibiotics belonging to the tetracycline family against P. aeruginosa. There was synergy between doxycycline and CP762 for most isolates that produced pyoverdine. Using a novel approach, this study showed different levels of synergy using a pre-made iron-doxycycline complex, highlighting iron-mediated inactivation of antibiotics. The findings indicate that CP762 has potential for use as an adjunct to doxycycline through sequestration of iron, thereby conferring synergistic activity to an antibiotic that would not normally be considered a therapeutic option against P. aeruginosa.
Regarding the lactoferrin/doxycycline antibacterial synergy hypothesis I did a literature search and in 1999 Alkawash et al appeared to find exactly that and by a large margin too, of 32 to 64 fold against B. cepacia.2
This was sufficient to overcome the bacterial resistance to the antibiotic at non-cytotoxic levels, thus potentially improving outcomes for the patients and increasing available treatment options.
MIC: Minimum Inhibitory Concentration of the drug required to inhibit or kill the pathogen.
MBC: Minimum Bactericidal Concentration. Less is better.
Abstract
The presence of lactoferrin at the concentration found in cystic fibrosis (CF) sputum (0.9 g/L) reduced MICs and MBCs of doxycycline for Burkholderia cepacia and Pseudomonas aeruginosa strains. MICs for B. cepacia fell by 32- to 64-fold, from highly resistant to clinically achievable values. Rifampicin MICs for B. cepacia strains were reduced by lactoferrin and for some strains MBCs were reduced. These findings suggest new therapeutic approaches to infections and question the relevance of standard sensitivity tests for CF pathogens. Addition of lactoferrin to media for the routine sensitivity testing of CF isolates might give more relevant results.
Our study arose in part from the clinical impression of benefit conferred by doxycycline therapy in CF patients with respiratory infection by P. aeruginosa. The in-vitro effect of lactoferrin in reducing the MICs of doxycycline for P. aeruginosa points to a possible basis for the impression, although MICs of our strains in the presence of lactoferrin were still quite high. The far greater effect on MICs of doxycycline for B. cepacia, an important pathogen for adult CF patients, points to a potentially important new approach to treatment and the necessity for a controlled clinical trial.
Doxycycline as an antiviral
In 2015, Wu et al investigated its effects in vitro on the replication of vesicular stomatitis virus.3
Of particular note here for also treating long covid/vaccine sequalae is that doxycycline (Dox) acts as both an antiviral and an anticancer therapeutic agent by the induction of expression of the key tumor suppressor p53.
Transfection induced disruption of p53 & BRCA and the potential consequences are discussed in these Substacks. The first paper was later retracted, but the grounds for retraction aren’t clear and seem unrelated to the experimental method or its findings:
Doxycycline (Dox) is a tetracycline derivative with broad-spectrum antimicrobial activities that is used as an effector substance in inducible gene-expression systems. We investigated the antiviral activity of Dox against vesicular stomatitis virus (VSV) infection in cultured H1299 cells. Dox at concentrations of 1.0–2.0 μg ml−1 significantly inhibited VSV replication and the VSV-induced cytopathic effect in dose-dependent manners, suggesting that Dox may have broader activity in inhibiting viral replication, in addition to its well-defined bacteriostatic activity. Dox exerted its antiviral effect at the early–mid stage of VSV infection, suggesting that it did not interfere with VSV infectivity, adsorption, or entry into target cells. These results indicate that Dox can inhibit VSV infection and may therefore have potential applications for the treatment of viral infections.
Dox is currently the preferred effector substance used in inducible gene-expression systems for eukaryotic cell biology research. In this system, the transcription of genes of interest is reversibly turned on (Tet-on) or off (Tet-off) in response to Dox stimulation (Gossen and Bujard ; Zhou et al. ).
The tumor suppressor p53 is a well-known protein that plays essential role in regulating cellular activities including apoptosis, cell cycle arrest, differentiation and antitumor response (Yonish-Rouach et al. ; Shaw et al. ; el-Deiry et al. ). Recent studies indicated a role for p53 in the host antiviral response to a range of viruses including vesicular stomatitis virus (VSV) (Takaoka et al. ), influenza virus (Turpin et al. ) and other viruses (Rivas, Aaronson and Munoz-Fontela ). To extend the antiviral spectrum of p53, we generated a stable cell line with inducible p53 expression basing on the Tet-on system, using Dox as the effector substance. We tested the inhibitory effect of Dox-induced p53 on the replication of VSV and Japanese encephalitis virus (JEV) in this stable cell line. Unexpectedly, Dox itself also inhibited VSV and JEV replication in this stable cell line with Dox-inducible p53 expression.
Figure 1
Generation of a stable H1299 cell line with Dox-inducible p53 expression. p53-null H1299 cells were transfected with plasmids pCMV-Tet3G and pTRE-p53 and selected with G418 and puromycin, respectively. The generated cell line (p53-Tet H1299) was treated with 1 μg ml−1 Dox and incubated for 24 h. (A) Dox-induced p53 expression was determined by western blot, (B) and localized by immunofluorescence analysis (green). Cells were stained for DNA with 4′-6-diamidino-2-phenylindole (blue). (C) The expression of p53 target genes (TLR3, p21 and BAX) was detected by qRT–PCR analysis. +Dox, cells treated with Dox; −Dox, cells mock-treated with equivalent H2O. **P < 0.01 between +Dox and −Dox groups analyzed by the Student's t-test.
Lower is better:
Figure 2
Dox inhibited the replication of VSV and JEV. p53-Tet H1299 and H1299 cells were infected with VSV at a MOI of 0.01 (A) or JEV at a MOI of 0.1 (B), respectively. After adsorption for 1 h, the cells were treated with Dox at 1 μg ml−1 and incubated for 24 h. Virus titers in the culture supernatants were determined by TCID50 assay. (C) Analysis of Dox cytotoxicity. H1299 cells were treated with the indicated concentrations of Dox for 24 h and cytotoxicity of Dox was analyzed using the cell proliferation reagent WST-1. Cell viability was calculated and plotted. The 50% cytotoxic concentration (CC50) was analyzed using non-linear regression curve fit analysis in the GraphPad Prism software. Data given as mean ± SD from three independent experiments. *P < 0.05 compared with cells mock-treated with equivalent H2O. +Dox, cells treated with Dox; −Dox, cells mock-treated with equivalent H2O.
Figure 3
Dox inhibited VSV replication in a dose-dependent manner. H1299 cells (A) and A549 cells (C) were infected with VSV at 0.01 MOI and treated with Dox at the indicated concentrations. Virus titers in the supernatants were determined by TCID50 assay 24 h post-infection. (B) The reduction of virus titer in H1299 cells from Fig. 3A was calculated and plotted using non-linear regression curve fit in the GraphPad Prism software. The x-axis is in a base 2 logarithmic scale. (D) H1299 cells infected with VSV and treated with Dox at the indicated concentrations were visualized under a light microscope 24 h post-infection. (E) H1299 cells infected with VSV and treated with Dox at the indicated concentrations were analyzed by cytotoxicity assay 24 h post-infection. Cell viability was calculated and plotted. Data are given as mean ± SD from three independent experiments. **P < 0.01 compared with cells mock-treated with equivalent H2O (0 μg ml−1 Dox). (F) Inhibitory effect of Dox on VSV-induced plaque formation. BHK-21 cells in six-well plate were infected with 10, 60 or 360 PFU VSV for 1h, then overlay with 1% agarose gel containing 1.0 μg ml−1 Dox. After incubating cultures for 4 days, monolayers were fixed with 4% formaldehyde solution for 30 min and stained with 1% (w/v) crystal violet solution.
Dox inhibits VSV replication in vitro at the early–mid stage of viral infection
H1299 cells were treated with Dox at a concentration of 2.0 μg ml−1 at different time points before VSV infection and during VSV infection (Fig. 4A). The viral titers in the supernatants of each group were analyzed to determine the stage at which Dox inhibited VSV replication. As shown in Fig. 4B, no significant inhibitory effect on VSV replication was observed in the groups treated with Dox before viral infection (Pre 2, 5 and 10 h), or during viral adsorption period (Ad 0 h), suggesting that Dox did not interfere with VSV infectivity, adsorption, or entry into target cells. However, Dox administered at the early stage (Post 1 and 2 h) or the mid stage (Post 5 h) of VSV infection showed a significant inhibitory effect on VSV replication. The most effective inhibition on VSV replication was observed at 2 h post-infection (Post 2 h). Dox administered at the late stage of VSV infection (Post 10 h) exerted no antiviral activity. Taken together, these results suggest that Dox inhibited VSV replication in vitro at the early–mid stage of VSV infection.
Figure 4
Dox inhibited VSV replication at the early–mid stage of viral infection. (A) Schematic representation of experimental design. H1299 cells were treated with Dox at a concentration of 2.0 μg ml−1 at the indicated times. (B) Virus titers were determined 24 h post-infection by TCID50 assay and plotted. Data are given as mean ± SD from three independent experiments. *P < 0.05 and **P < 0.01 compared with cells mock-treated with equivalent H2O (0 μg ml−1 Dox).
DISCUSSION
Dox is a semi-synthetic structural isomer of the tetracycline family and one of the most widely used broad-spectrum antibiotics in clinical practice (Bobbio 1969; Misra et al. 1969; Mita et al. 1977; Caton and Ryan 2011). It exhibits intracellular penetration and bacteriostatic activity in many bacteria by interfering with protein synthesis (Semenkov Yu et al. 1982). The antiviral activity of Dox was first demonstrated by a significant reduction in retrovirus titers in cultured cells treated with Dox (Sturtz 1998). Dox was subsequently shown to inhibit DNV infection (Yang et al. 2007). These observations suggest that Dox may have broader activity in inhibiting viral replication, in addition to its well-defined bacteriostatic activity.
We ruled out the possibility that the inhibitory effect of Dox on VSV and JEV replication was the result of Dox cytotoxicity.This result was consistent with previous observations that 0.01–2.0 μg ml−1 Dox had no obvious cytotoxicity in vitro (Kees et al. 1990; Beringer et al. 2012; Rothan et al. 2014). Taken together, our results demonstrated a new role for Dox in inhibiting the replication of VSV, further reinforcing the concept that Dox can inhibit viral infection. However, Dox showed no effect on virus titers in mice infected with virulent influenza H3N2 virus (Ng et al. 2012). The antiviral activity of Dox may therefore depend on the virus species. Further studies are needed to clarify the antiviral spectrum of Dox.
The mechanisms whereby Dox inhibits viral replication are poorly understood. Dox inhibits the propagation of type 2 DNV, probably by disrupting conformational changes of the DNV E protein that is required for virus entry (Yang et al. 2007). In addition to inhibiting viral entry, Dox has also been shown to inhibit viral NS2B-NS3 serine protease and post-infection replication of DNV (Rothan et al. 2014). In the current study, Dox exerted its antiviral activity at the early–mid stage of VSV infection, suggesting that it did not affect VSV entry into target cells, or virus infectivity or adsorption. However, further studies are needed to determine the exact mechanisms whereby Dox inhibits VSV replication.
Dox is the preferred effector substance used in the inducible gene-expression system for eukaryote cell biology research. However, this study unexpectedly demonstrated the anti-VSV activity of Dox in the stable cell line p53-Tet H1299 with Dox-inducible p53 expression.Dox also shows a variety of bioactivities including inhibition of matrix metalloproteases (Dursun et al. 2001), reduction of inflammation (Angadi 2012) and cell cycle arrest at G1 phase (Duivenvoorden, Hirte and Singh 1997). Careful attention should therefore be paid to Dox-inducible gene expression to avoid misinterpretation of experimental results.
In conclusion, inhibitory effect of of Dox on VSV replication was discovered unexpectedly when study antiviral function of p53 in p53-Tet H1299 cells. The anti-VSV activity of Dox further evaluated in H1299, A549 and BHK-21 cells indicated that Dox significantly inhibited VSV replication and the VSV-induced CPE in dose-dependent manners. Dox exerted its antiviral activity in vitro at the early–mid stage of VSV infection, suggesting that it did not affect VSV entry into target cells, or virus infectivity or adsorption. These results provide evidence to support the ability of Dox to inhibit viral infection, and suggest the potential application of Dox for the treatment of viral infections.
More on doxycycline induction of p53 expression. Son et al (2009) conducted an in vitro investigation and concluded that the tumor suppressive effects on pancreatic cancer cells were due to the activation of proapoptotic genes, inhibiting IL-8 expression, and suppressing antiapoptotic genes.4
“Tumor protein P53, also known as p53, cellular tumor antigen p53 (UniProt name), the Guardian of the Genome,[5]phosphoprotein p53, tumor suppressor p53, antigen NY-CO-13, or transformation-related protein 53 (TRP53), is any isoform of a protein encoded by homologous genes in various organisms, such as TP53 (humans) and Trp53 (mice). The p53 proteins (originally thought to be, and often spoken of as, a single protein) are crucial in vertebrates, where they prevent cancer formation. As such, p53 has been described as "the guardian of the genome" because of its role in conserving stability by preventing genome mutation. Hence TP53 is classified as a tumor suppressor gene”
“p21Cip1 (alternatively p21Waf1), also known as cyclin-dependent kinase inhibitor 1 or CDK-interacting protein 1, is a cyclin-dependent kinase inhibitor (CKI) that is capable of inhibiting all cyclin/CDK complexes, though is primarily associated with inhibition of CDK2. p21 represents a major target of p53 activity and thus is associated with linking DNA damage to cell cycle arrest. This protein is encoded by the CDKN1A gene located on chromosome 6 (6p21.2) in humans…Studies of p53 dependent cell cycle arrest in response to DNA damage identified p21 as the primary mediator of downstream cell cycle arrest”.
“The Fas receptor, also known as Fas, FasR, apoptosis antigen 1 (APO-1 or APT), cluster of differentiation 95 (CD95) or tumor necrosis factor receptor superfamily member 6 (TNFRSF6), is a protein that in humans is encoded by the FAS gene. Fas was first identified using a monoclonal antibody generated by immunizing mice with the FS-7 cell line. Thus, the name Fas is derived from FS-7-associated surface antigen.[
The Fas receptor is a death receptor on the surface of cells that leads to programmed cell death (apoptosis) if it binds its ligand, Fas ligand (FasL). It is one of two apoptosis pathways, the other being the mitochondrial pathway.”
Background: Tetracyclines such as doxycycline are reported to possess cytotoxic activity against mammalian tumor cells, but the mechanism of their effects on cell proliferation remains unclear.
Materials and methods: The antitumor effect of doxycycline was investigated in human pancreatic cancer cell line, PANC-1. We also investigated the effect of doxycycline on expression of a potent proangiogenic factor, interleukin (IL)-8.
Results: In excess of 20 microg/ml, cytotoxic effects of doxycycline were accompanied by G(1)-S cell cycle arrest and DNA fragmentation in PANC-1 cells. Doxycycline consistently activated transcription of p53, p21 and Fas/FasL-cascade-related genes, while reducing the expression of Bcl-xL and Mcl-1. Doxycycline (5 microg/ml) below the cytotoxic level suppressed endogenous and paclitaxel-induced IL-8 expression. In the mouse xenograft model, doxycycline treatment was shown to suppress tumor growth by 80%.
Conclusion: These data suggest that doxycycline exerts its antitumor effect by activating proapoptotic genes, inhibiting IL-8 expression, and suppressing antiapoptotic genes.
Doxycycline induces the expression of p53 and its downstream target p21 in PANC-1 cells. The findings of the previous set of experiments suggested that doxycycline could induce cell cycle arrest and apoptosis in PANC-1 cells. One possible mechanism by which doxycycline participates in cell cycle arrest and apoptosis is the involvement of the tumor suppressor gene p53. The activation of p53 probably causes cell cycle arrest through the transcriptional up-regulation of its downstream targets including p21, and/or a transcription-independent apoptosis. Therefore, we investigated the effect of doxycycline on the levels of p53 and p21. PANC-1 cells were treated with doxycycline for 48 h. Following treatment, cells were harvested and total RNA was prepared for Northern blot analysis. As shown in Figure 3A, p53 mRNA level was induced after exposure of PANC-1 cells to doxycycline. This induction was observed at as low as 15 μg/ml of doxycycline and increased at higher doses in a dose-dependent manner. Like p53 mRNA up-regulation, p21 mRNA was increased in excess of 15 μg/ml doxycycline (Figure 3A). Up-regulation of p53 mRNA appeared at as early as 24 h after exposure to 20 μg/ml doxycycline (Figure 3B), in agreement with G1-S arrest determined by FACS analysis. Furthermore, 40 μg/ml doxycycline treatment resulted in earlier p53 induction at 12 h (Figure 3B), followed by detectable p21 mRNA induction at 24 h. These transcriptional activations of p53 and p21 genes support the assumption of doxycycline-mediated G1-S cell cycle arrest and subsequent induction of apoptosis.
Figure 1.
Growth inhibition by doxycycline in PANC-1 cells. A, PANC-1 cells were incubated with different concentrations of doxycycline. Cell growth was inhibited by doxycycline in a dose-dependent manner. Bars, SD. B, Demonstration of apoptosis by DNA fragmentation. Lane M, 1 kb DNA ladder; lane 1, DNA from control cells incubated with PBS (as shown 0 μg/ml) for 48 h; Lanes 2 to 6, DNA from cells cultured with increasing concentrations of doxycycline for 48 h. Note laddering in lanes 5 and 6, consistent with fragmentation of DNA.
A variety of chemotherapeutic agents can cause some degree of DNA damage, resulting in a rapid up-regulation of p53 protein levels by a posttranscriptional mechanism. The outcome of the activation of p53 is either apoptosis or cell cycle arrest, depending on the level of p53 expression: p53 at high levels promotes cell death by a transcriptionally independent mechanism, whereas lower levels initiate a transcriptionally controlled inhibition of cell cycle progress. We have previously reported that doxycycline-induced apoptosis is a nuclear factor (NF)-κB- and p53-dependent in colon cancer cell line HCT116. Similarly, simultaneous activation of p53 and p21 as well as cell cycle arrest were observed in PANC-1 cells, suggesting that a p53-dependent mechanism is involved in doxycycline-mediated apoptosis.
In conclusion, doxycycline exhibited cytotoxic effects in pancreatic cancer PANC-1 cells and a mouse xenograft model. Doxycycline-mediated apoptosis seems to be involved in several apoptotic signal pathways, including caspase activation and the p53 signal pathways. Additionally, low-dose doxycycline blocked paclitaxel-induced IL-8 expression. Further studies are warranted to determine the potential efficacy of doxycycline as an anticancer agent as well as an enhancer of conventional chemotherapeutic agents.
Figure 6.
Effects of doxycycline on tumor growth in a nude mouse xenograft model. Overview of mice implanted with placebo (A) and doxycycline pellet (B) seven weeks after doxycycline therapy. C, Changes in tumor volume. D, Tumor xenograft weight at seven weeks after doxycycline therapy. There were significant differences in tumor volume and weight between the groups.
In 2020, Mosquera-Sulbaran and Hernández-Fonseca published a review on the use of tetracycline as an anti COVID-19 therapeutic:5
Abstract
Tetracyclines have been used to treat many bacterial infections. The use of these antibiotics for the treatment of viral diseases dates to the 1960s to 1970s. Over the decades, the effect of tetracyclines on the pathogenesis of viral infections has been demonstrated both clinically and experimentally. Tetracyclines can act on viral infections either through their antibacterial properties or through direct antiviral action. This review focuses on clinical and experimental data that support the use of tetracycline in treating viral infections and highlights an important approach to slowing disease progression during viral infections. Tetracycline treatment might represent a strategy for eliminating the infection or inhibiting the progression of COVID-19.
Dox has also shown antiviral activity. Recent studies have demonstrated the importance of the p53 protein in the host antiviral response against several viruses, including vesicular stomatitis virus (VSV) and influenza virus.The ability of Dox to induce p53 probably extends the antiviral effect of this antibiotic. In this regard, the inhibition of p53 expression by JEV in H1299 cell cultures was inhibited by Dox. Dox also inhibited the cytopathic effect and viral replication in H1299 cell cultures infected by VSV. This antibiotic significantly reduced the cytopathic effect and viral replication in Marc-145 cells infected by swine respiratory syndrome virus. Dox also reduced the severity of acute lung injury in mice infected with influenza H3N2 virus. The antiviral effect of Dox against flaviviruses has also been reported. In vitro studies have shown that Dox prevents the entry and replication of chikungunya virus in Vero cell cultures. Dox also inhibited the serine protease of dengue virus (DENV NS2B-NS3pro), inhibiting the entry and replication of four dengue serotypes (DENV2, DENV4, DENV1 and DENV3) in Vero cell cultures. In addition to this mechanism, Dox, through its tetracyclic rings, can alter the conformation of the dengue virus envelope protein and block the entry of this virus into the cells (Fig. 3).
Suppression of the p38 MAPK signalling pathway is another key anti-cancer mechanism, as well as reducing high inflammatory cytokine levels.
And matrix metalloproteinase (MMPs) can be particularly damaging to the blood brain barrier, the cardiovascular system and joints, as with rheumatoid arthritis (RA):
Fig. 3 Effect of tetracyclines on viral pathogenesis. Tetracyclines can affect the viral life cycle by indirect and direct mechanisms. Indirect effects are based on anti-apoptotic, anti-metalloproteinase (MMPs), anti-oxidative stress, and anti-inflammatory properties of these compounds. Different mechanisms of antiviral activity of tetracyclines have been reported. Overall, the properties of tetracycline by acting on viral pathogenesis leads to decreased entry of the virus into the host cell, decreasing its replication and cellular damage. MIF, macrophage inhibitory factor
They, quite correctly, caution against the risk of viral resistance in addition to bacterial resistance due to misuse - ie taking as a prophylactic if you are not infected or a cancer patient.
Viral resistance to tetracycline could be a problem in tetracycline treatment. Antibiotic resistance occurs when germs like bacteria and fungi develop the ability to defeat the drugs designed to kill them. Tetracycline resistance is widespread among Gram-positive and Gram-negative bacteria and can be the result of pumping the drug out of the cell before it reaches its site of action (efflux), protection of the ribosomal binding site, which decreases drug binding, or changes in the permeability of the cell envelope that reduce drug uptake. Viruses are not living particles; they need host cells to replicate. The antiviral effect of tetracyclines is indirect and involves several mechanisms that the viruses use to enter and replicate cells. The blockage of protein synthesis in the cell by tetracyclines might inhibit virion formation. Thus, it is unlikely that the mechanisms of antibiotic resistance used by viruses will be similar to those used by bacteria…
COVID-19 and tetracyclines
…Tetracyclines (e.g., tetracycline, halogenated tetracyclines, doxycycline, and minocycline) are highly lipophilic antibiotics that are known to chelate zinc compounds on matrix metalloproteinases (MMPs). Several functions of coronavirus are associated with the host MMPs complex, including replication. Therefore, the zinc-chelating properties of tetracyclines may also aid in inhibiting SARS-CoV-2 infections in humans, limiting their ability to replicate within the host. Tetracyclines might also be able to inhibit RNA replication of positive-sense single-stranded RNA viruses, like SARS-CoV-2, and they can downregulate the NF-κB pathway, decreasing the levels of inflammatory cytokines such as TNF‐α, IL‐1β, and IL‐6 induced by SARS-CoV-2. In addition, tetracyclines stand out as potential inhibitors of the main protease of SARS-CoV-2 (Fig. 4). In this context, various investigators have suggested the use of tetracyclines as potential therapeutic agents for the treatment of COVID-19.
Fig. 4. Potential effects of tetracyclines on SARS-CoV-2. Several properties of tetracyclines can potentially decrease the ability of SARS-CoV-2 to enter and reproduce in the cell
A interesting point regarding apoptosis is that a cell invaded by a virus needs to be eliminated by the organism as an infected cell, using various mechanisms, including the antiviral immune response, but the virus requires a living cell for reproduction, and in the late period of infection can use apoptosis as a means of breaking the cell and increasing the spread of the virus. Therefore, the antiviral effect of tetracyclines on apoptosis may be related to the spread of infection. When tetracyclines induce apoptosis, they can favor the dissemination of the virus, but at the same time decrease its replication, but when tetracyclines block apoptosis, they can decrease spread of the virus. It has been reported that SARS-CoV-2 induces apoptosis in infected cells, and the effect on this process of tetracycline remains unknown.
Although several studies have shown that tetracyclines have antiviral and anti-inflammatory effects, all of them have been in vitro studies using experimental models. There are still no final results of clinical studies demonstrating the efficacy of these antibiotics for treatment of patients with COVID-19.
There has been only one phase 2 study in which doxycycline was used together with ivermectin for the treatment of COVID-19 patients.However, the potential of tetracyclines as an antiviral drug is high and requires further clinical studies. It is important to develop antiviral treatments for COVID-19 that can be administered early in infection in order to avoid the organ damage caused by the virus and allow the patient to produce a strong immune response against the virus. Therapeutic interventions in patients with severe organ damage caused by the virus or by an excessive immune response may have little effect on eliminating the virus or the recovery of the patient when antiviral strategies are given at later times.
I was unable to find the results of the study referenced above, other than the trial details (72 participants):
Efficacy and Safety of Ivermectin and Doxycycline in Combination or IVE Alone in Patients With COVID-19 Infection (2020)
But in 2021, Mahmud conducted a similar randomized control trial, this time with 400 participants and found a significant reduction in time to clear and severity of symptoms:6
Abstract
Objective: We evaluated whether ivermectin combined with doxycycline reduced the clinical recovery time in adults with COVID-19 infection.
Methods: This was a randomized, blinded, placebo-controlled trial in patients with mild-to-moderate COVID-19 symptoms randomly assigned to treatment (n = 200) and placebo (n = 200) groups. The primary outcome was duration from treatment to clinical recovery. Secondary outcomes were disease progression and persistent COVID-19 positivity by RT-PCR.
Results: Among 556 screened patients, 400 were enrolled and 363 completed follow-up. The mean patient age was 40 years, and 59% were men. The median recovery time was 7 (4-10, treatment group) and 9 (5-12, placebo group) days (hazard ratio, 0.73; 95% confidence interval, 0.60-0.90). The number of patients with a ≤7-day recovery was 61% (treatment group) and 44% (placebo groups) (hazard ratio, 0.06; 95% confidence interval, 0.04-0.09). The proportion of patients who remained RT-PCR positive on day 14 and whose disease did not progress was significantly lower in the treatment group than in the placebo group.
Conclusions: Patients with mild-to-moderate COVID-19 infection treated with ivermectin plus doxycycline recovered earlier, were less likely to progress to more serious disease, and were more likely to be COVID-19 negative by RT-PCR on day 14.
Declaration of conflicting interest: The authors declare that there is no conflict of interest. Popular Pharmaceuticals Limited, Bangladesh provided ivermectin, doxycycline, and placebo. The company was not involved in the planning or design of the study and had no role in the collection, analysis, or interpretation of the data.
Numerous clinical trials worldwide have explored the efficacy of existing medicines against COVID-19, including various antiviral and immunomodulatory drugs. Australian researchers from Monash University established the efficacy of ivermectin, a broad-spectrum antiviral drug, against severe acute respiratory syndrome-related coronavirus (SARS-CoV-2) in in vitro studies; viral replication was stopped within 24 to 48 hours. Tetracycline, a widely available and well-tolerated antibiotic useful against atypical infections, has known anti-inflammatory effects and, along with its synthetic derivative, doxycycline, has been shown by Mohit et al. to potentially be effective against COVID-19. The antiviral and anti-inflammatory properties of ivermectin combined with doxycycline may be beneficial in the treatment of COVID 19. Given that these two drugs have different modes of action, their synergistic effects may contain viral infection by targeting different sites of disease pathogenesis. In the present study, we sought to determine the efficacy of this combination in patients with mild-to-moderate COVID-19 symptoms.
Study interventions
The treatment group received a single dose of ivermectin 12 mg and doxycycline 100 mg, twice daily for 5 days, in addition to standard of care. Standard of care included administration of paracetamol, antihistamines, cough suppressants, vitamins, oxygen therapy according to indication and need, low molecular weight heparin according to indication, appropriate other broad-spectrum antibiotics, remdesivir injection, other antiviral drugs, and other drugs for associated comorbid conditions.
The placebo group received placebo in addition to standard of care.
Figure 2. Time-to-recovery in the treatment and control groups, with and without censored data. Hazard ratio (95% confidence interval): 0.73 (0.60–0.90); P = 0.003.
Figure 3. Post-hoc analysis of time-to-recovery among the subgroups. Data are presented as hazard ratios and 95% confidence intervals.
Safety outcomes
Among the 400 included patients, adverse drug reactions occurred in 9 patients (2.5%); of these, 2 patients discontinued intervention owing to erosive esophagitis. Non-ulcer dyspepsia developed in seven (1.75%) patients (Appendix 1). Three patients in the placebo group died; these patients had a higher mean age than those who survived (63 years vs. 39 years) (Appendix 2) and they died 8, 22, and 28 days after randomization of respiratory failure due to COVID-19-related pneumonia.
We were encouraged by the in vitro research findings of researchers at Monash University, Australia;6 the study revealed that ivermectin could reduce viral replication within 24 to 48 hours of treatment. There was a >5000-fold reduction in viral RNA with 5 μM ivermectin in cell culture, equating to a >99% reduction in viral RNA. Schmith et al. stated, “The concentration resulting in 50% inhibition (IC50; 2 µM) was >35× higher than the maximum plasma concentration (Cmax) after oral administration of the approved dose of ivermectin 200 μg/kg when given fasted,” and predicted that with the oral dose of ivermectin 200 μg/kg, lung concentrations would be approximately one-fourth of the IC50. The safety of higher doses has not been evaluated in humans. Therefore, we used the conventional dose of ivermectin in the present study.
Doxycycline, which may be used for the treatment of atypical bacterial pneumonia and community-acquired pneumonia, exerts an anti-inflammatory effect mediated by chelating zinc compounds on matrix metalloproteinases (MMPs) in mammalian cells. Doxycycline also has antiviral activity, especially against dengue virus and Chikungunya virus. A previous in vitro study showed that murine coronaviruses rely on MMPs for cell fusion and viral replication. The pathologic features of COVID-19 closely resemble those of other SARS-CoV infections, where MMPs play an important role in disease pathogenesis. Therefore, doxycycline may potentially be effective for the treatment of COVID-19 infection.
Ivermectin and doxycycline were co-administered in the treatment group because their synergistic action may increase the likelihood of efficacy in the treatment of COVID-19. We did not observe known drug–drug interactions between ivermectin and doxycycline; therefore, no drug dosage modification was required in the present study.
The effective dose of ivermectin required to reach IC50 at a pulmonary level is considerably higher than that used in this study. However, evaluation at higher doses requires detailed safety analysis, which was not within the scope of the present analysis. Therefore, approved dosing regimens of ivermectin and doxycycline were used in this study.
We used approved dosing regimens for both drugs and noted very few adverse reactions. Furthermore, most reactions were consistent with those associated with doxycycline, which supports our assumption that the combination treatment was safe to use in the study population. The death rate observed (three of 200 patients in the placebo group; 1.5%) was consistent with the COVID-19 mortality rate in Bangladesh (1.4%)
Our study was performed at a single center over a short period. Therefore, our findings need to be carefully interpreted. Furthermore, a priori sample size calculations were not performed, limiting the strength of our findings. We could not test for viral load, and therefore could not directly assess viral clearance. It remains unclear whether the early reductions in viral load we observed should be verified by repeated RT-PCR testing during follow-up. Our testing facilities did not allow us to test patients repeatedly. Finally, we evaluated the combination of doxycycline and ivermectin but did not determine their individual effects. Therefore, further studies are needed to clarify the results observed in the present study.
Conclusions
Adult patients with mild-to-moderate COVID-19 infection treated with ivermectin combined with doxycycline recovered earlier than those receiving placebo, were less likely to progress to a serious disease, and were more likely to test negative for COVID-19 at the end of the treatment period. Although additional research on the effects of ivermectin combined with doxycycline is warranted, the safety and efficacy of this combination are favorable compared with current standard of care.
Iron chelation and inhibition of tumors and metastasis
There are many papers on this, I will highlight three from 2003, 2013 and 2021.
Buss et al (2003) recognized the potential of using iron chelation in cancer therapy and their possible synergistic effects (paywalled):7
Abstract
This review focuses on advances and strategies in the use of iron chelators as anti-tumor therapies. Although the development of iron chelators for human disease has focused primarily on their use in the treatment of secondary iron overload, chelators may also be useful anti-tumor agents. They can deplete iron or cause oxidative stress in the tumor due to redox perturbations in its environment. Iron chelators have been tested for their anti-tumor activity in cell culture experiments, animal models and human clinical trials. Largely for pragmatic reasons, clinical studies of the anti-tumor activity of iron chelators have generally focused on desferrioxamine (DFO), a drug approved for the treatment of iron overload. These studies have shown that DFO can retard tumor growth in many different experimental contexts. However, the activity of DFO is modest, and advances in the use of chelators as anti-cancer agents will require the development of new chelators based on new paradigms. Examples of iron chelators that have shown promising anti-tumor activity (in various stages of development) include heterocyclic carboxaldehyde thiosemicarbazones, analogs of pyridoxal isonicotinoyl hydrazone, tachpyridine, O-trensox, desferrithiocin, and other natural and synthetic chelators. Apart from their use as single agents, chelators may also synergize with other anti-cancer therapies. The development of chelators as anticancer agents is largely an unexplored field, but one with extraordinary potential to impact human cancer.
From 2013, Richardson et al review how the iron chelator DFO can inhibit key signalling pathways which induce epithelial mesenchymal transition (EMT) in pancreatic cancer and other tumors (paywalled).8
EMT: “The epithelial–mesenchymal transition (EMT) is a process by which epithelial cells lose their cell polarity and cell–cell adhesion, and gain migratory and invasive properties to become mesenchymal stem cells; these are multipotent stromal cells that can differentiate into a variety of cell types. EMT is essential for numerous developmental processes including mesoderm formation and neural tube formation. EMT has also been shown to occur in wound healing, in organ fibrosis and in the initiation of metastasis in cancer progression.”
Pancreatic cancer is the fourth leading cause of cancer-related deaths worldwide in both men and women. It presents late with non-specific symptoms, which makes it difficult to diagnose until the cancer has progressed and metastasized. Metastasis is facilitated by the epithelial-to-mes-enchymal transition (EMT), which is promoted via the oncogenic transforming growth factor beta (TGFβ), Wnt, and nuclear factor kappa B (NFκB) signaling pathways. However, recent studies have demonstrated that the EMT can be inhibited by novel anti-cancer agents known as thiosemicarbazone iron chelators. These novel agents also up-regulate the metastasis suppressor, N-myc downstream regulated gene 1 (NDRG1), which can restore normal signaling to the cell and suppresses metastasis via inhibition of the EMT. Through the ability of iron chelators to up-regulate NDRG1 expression and affect multiple molecular targets, these agents have the potential to maintain the epithelial phenotype of cancer cells and may lead to improved survival rates for patients with late-stage disease.
Our final paper from 2021 by Morales and Xue reviews the targeting of iron metabolism in cancer therapy.9
Its a broad subject worthy of reading in its entirety, but for brevity I will focus on key aspects:
Abstract
Iron is a critical component of many cellular functions including DNA replication and repair, and it is essential for cell vitality. As an essential element, iron is critical for maintaining human health. However, excess iron can be highly toxic, resulting in oxidative DNA damage. Many studies have observed significant associations between iron and cancer, and the association appears to be more than just coincidental. The chief characteristic of cancers, hyper-proliferation, makes them even more dependent on iron than normal cells. Cancer therapeutics are becoming as diverse as the disease itself. Targeting iron metabolism in cancer cells is an emerging, formidable field of therapeutics. It is a strategy that is highly diverse with regard to specific targets and the various ways to reach them. This review will discuss the importance of iron metabolism in cancer and highlight the ways in which it is being explored as the medicine of tomorrow.
Keywords: Iron metabolism, Cancer, Therapy, Chelation, Ferroptosis
Introduction
Iron is essential for cell vitality. It is found in proteins that perform a variety of functions including biomolecule synthesis, oxygen transport and homeostasis, and respiration. Iron is a critical component of many proteins involved in nucleic acid metabolism and repair, as well as cell cycle progression. Because iron is an integral component of anatomy and physiology and its bioavailability is scarce, iron stores are tightly regulated within the body in order to ensure conservation and mitigate toxicity.
The oxidation-reduction (redox) ability of iron is at the heart of its importance as a handler of oxygen and electrons, but it is in this same role that it harbors its dangers. Iron is able to easily interconvert between the ferrous state (Iron [II]) and ferric state (Iron [III]) and may exist in a wider range of oxidation states. In cellular metabolism, iron largely draws its negative effects from the reduction of oxygen. Due to oxygen's atomic nature, its reduction must proceed in a stepwise fashion of individual electron additions and reactive intermediates. During this process, the Fenton reaction can occur between ferrous iron and hydrogen peroxide to generate the highly reactive hydroxyl radical. Oxygen reduction intermediates are known as reactive oxygen species (ROS) and have been linked to lipid, protein, nucleic acid, and various signaling pathway damage. As such, iron has become a key target of interest in the progression and treatment of diseases including cancer. This review will discuss aspects of the therapeutic potential of iron metabolism for cancer. First, we will present a brief overview of the role of iron in the body and discuss aspects of the therapeutic potential of iron for the treatment of cancer.
Iron absorption and recycling. Non-heme iron is absorbed into enterocytes by DMT-1 after reduction from Fe (III) to Fe(II) by DcytB. Iron is carried by chaperones such as PCBPs to sites for storage in ferritin or for functional usage in cellular proteins and metabolism. Iron can be exported through FPN and subsequently re-oxidized by HEPH to Fe (III). Most circulating iron is carried by Tf and delivered to various tissues via its receptor TfR1 through receptor-mediated endocytosis. Tf and ferric iron dissociate in the endosome, after which the ferric iron is reduced to ferrous iron by STEAP proteins and enters the cytosol. Circulating iron is mainly derived from phagocytosis in senescent red blood cells, a process mediated by macrophages. Iron loss from the body occurs regularly through tissue loss such as epithelial shedding and blood loss. DMT-1: divalent metal transporter 1; DcytB: duodenal cytochrome B; PCBP: poly(rC)-binding protein; FPN: ferroportin; HEPH: hephaestin; STEAP: six transmembrane epithelial antigen of the prostate; Tf: transferrin; TfR1: transferrin receptor 1.
Hepcidin is to iron regulation what insulin is to glucose regulation. Its a feedback mechanism that responds to changing cellular conditions such as hypoxia:
Major regulators of iron homeostasis include hepcidin, HIFs and IRP/IRE systems. [A] Hepcidin naturally limits the amount of iron efflux from cells and is inhibited by conditions such as anemia, hypoxia, increased testosterone, and increased erythropoiesis, while being upregulated by systemic iron overload and inflammation. [B] HIFs respond to low oxygen and iron levels and transcribe genes to help cells adapt to perceived environmental deficiencies for a more sustainable metabolism and long-term survival; this results in short- and long-term changes including increases in glycolysis, angiogenesis, iron supplies, and ultimately cell vitality. [C] IRPs control gene translation through binding of IREs on mRNA transcripts for iron metabolism-related proteins, either promoting translation through 3' UTR binding-dependent stabilization (e.g., TfR1, DMT1), or inhibiting translation through 5' UTR binding that results in eventual degradation (e.g., Ferritin, FPN). HIF: hypoxia-inducible factor; IRP: iron response protein; IRE: iron response element; mRNA: messenger RNA; UTR: untranslated region.
Iron and Cancer
As previously mentioned, there is an indisputable need for iron in the body. However, excess iron accumulation can be highly toxic. Dysregulated iron homeostasis due to hereditary and lifestyle factors may lead to increased risk of cancer. Cancer cells are characterized by rapid proliferation, which consequently demands a greater amount of iron and results in dysregulated levels of key proteins involved in iron metabolism.
Metabolic changes in iron-mediated carcinogenesis
Metabolic profiling of cancer cells enables scientists to better understand the implications of iron metabolism in cancer. An analysis of the metabolic profile of cholangiocarcinoma cells noted a strong shift towards iron retention in the cells, in agreement with the concept of iron dependence in cancer cells. The shift in cell metabolism towards iron accumulation also appears to play a role in the development of leukemia. Widespread dysregulation of iron metabolism in leukemia involves increased cellular iron import caused by overexpressed TfR1 and decreased iron export due to reduced expression of FPN. A study by Marques et al. noted a phenomenon of both iron acceptance by breast cancer cells, as well as iron donation by immune cells in the local tumor environment. One study shows that increased systemic hepcidin levels occurring as a result of a dysregulated hepcidin-FPN axis promotes breast cancer growth. Recently we found that ectopic hepcidin expression in colorectal cancer tissues is essential for maintaining cell proliferation due to the role of iron in both nucleotide synthesis and mitochondrial metabolism. Further implications of hepcidin-FPN axis regulation in cancer will be discussed later as a therapeutic avenue against cancer.
With the pattern of iron loading in mind, the question moves its focus towards what the cells do with the extra iron. As mentioned before, iron is essential for various cellular functions including the processes necessary for energy production and replication [See Iron Absorption and Recycling]. Accumulating intracellular iron affects the cell cycle; we and others have seen that iron likely binds to cyclin dependent kinase 1 (CDK1), a major mitotic component, and activates it to trigger pro-proliferative downstream signaling. CDK1 is overexpressed in colorectal cancer and predicts poor prognosis. Taken together, the body of studies on iron controlled CDK1 presents insight into one way that iron can manipulate the excessive growth characteristic of tumors. Furthermore, dysregulation of ribonucleotide reductase activity, essential for DNA synthesis, was found to result in carcinogenesis in vivo with particular concern on the M2 subunit overexpression being an accelerator of the malignant process.
Ni et al. reported that iron accumulated in the mitochondria through the upregulation of mitochondrial iron import proteins mitoferrin 1 and 2, and promoted a transformation of the cellular metabolism towards glycolysis known as the Warburg effect. The Warburg effect was discovered in the early 20th century and is thought to be an adaptation that simplifies energy production in exchange for fast-tracking synthesis of various biomolecules. An iron chelator prevented the Warburg effect, indicating the bona fide role of iron in cellular metabolic reprogramming. Mitochondrial iron can be repurposed through the process of mitophagy, a selective form of autophagy that removes damaged mitochondria. This process can help tumor cells survive and adapt, but it may not be entirely beneficial to cancer cells either. For example, we have recently shown that PTEN-induced kinase 1-dependent mitophagy is essential for suppressing colon tumor growth. This is consistent with a previous report showing that increases in degradation of iron-rich mitochondria by mitophagy in intestinal epithelial cells cause an excess of iron buildup in lysosomes, resulting in increases in ROS produced by the Fenton reaction. Subsequent addition of the lysosomal membrane permeabilization inducer chloroquine raises the pH of the lysosomes and compromises the integrity of the lysosomal membrane, resulting in an increased leakage of cathepsins, a group of proteases, into the cytosol and alters cellular antigen presentation to elicit cytotoxic T-cell immune responses. Thus, lysosomal iron accumulation can be exploited to induce anti-tumor immunity and restrict tumor growth.
In relation to oxidative stress, the accumulation of iron in cancer cells does bring the great dangers to the cells in addition to its benefits. In other words, cancer cells are no exception to the rules of iron; however, they develop ways to stay ahead. The glutamate-cystine antiporter, also known as system Xc-, plays a vital role in the antioxidant defense system of cells by supplying cystine to the cell; the amino acid regenerates glutathione, which terminates ROS through glutathione peroxidase 4 (GPX4). The system Xc- component xCT (light chain) encoded by the gene SLC7A11 is overexpressed in non-small cell lung cancer cells. High expression of SLC7A11 is associated with poorer outcomes and may contribute to metabolic reprogramming in tumors.
In summary, iron dysregulation may come in many forms through different sources, and cancer cells undergo alterations in their gene expressions to favor the import and retention of iron leading to larger supplies to sustain their rapid growth characteristics. These transformations may be regulated at various points and affect the biosynthetic and proliferative statuses of the cell. In order to avoid cellular damage from iron toxicity, cancer cells take advantage of antioxidant systems. Understanding iron toxicity, iron-dependency and addiction, and homeostatic disruption, including the various upstream controls and downstream effectors of the process, has become an increasingly popular area of study in terms of cancer treatment. The following section discusses the potential of targeting iron metabolism for therapeutic purposes in further detail.
Potential Cancer Therapeutic Targets and Biochemical Treatments
Due to the critical role of iron in cancer, targeting iron metabolism emerges as a novel therapeutic strategy in the treatment of cancer. Here we have chosen to highlight and discuss several promising strategies that are currently being explored in the world of cancer therapeutics (Figure 3).
Figure 3
Potential therapeutic pathways in cancer targeting abnormal iron metabolism. [A]Iron chelators limit the available amount of iron in tumor cells. Chelators and other drugs can be delivered through a TfR1-mediated drug delivery. [B] Inhibiting TfR1 by antibodies and gene silencing can lower iron import, depriving the cell of its desired iron content. [C] Inhibition of HIFs and their target genes can ultimately limit the amount of iron available for cells and hinder the ability of cancer cells to proliferate. [D] Inhibition of hepcidin-FPN axis can increase cellular iron export depriving the cell of iron. [E] Inhibition of cellular antioxidant defenses such as system xC- and GPX4 renders the cell prone to ROS accumulation from iron metabolism, leading to lipid peroxidation and ferroptosis. TfR1: transferrin receptor; HIFs: hypoxia-inducible factors; FPN: ferroportin; xC-: cystine-glutamate antiporter; GPX4: glutathione peroxidase 4; ROS: reactive oxygen species.
Iron Chelation
Iron chelation is a developing strategy aimed at sequestering iron from usage in tumor cells (Figure 3). Iron chelators had been used extensively to treat disorders of iron-overload in order to help patients evade the effects of iron toxicity. Deferoxamine (DFO), deferiprione (DFP), and deferasirox (DFX) are three chelators commonly used in clinical settings. However, there are varying degrees of toxicity with them as well. DFO treatment of breast cancer cell lines MCF-7 and MDA-MB-231 resulted in significant reduction of the intracellular iron supply and decreased cellular regeneration and survival. Another study of DFO using breast cancer cell lines showed strong results in combining chelation with radiation treatment, which resulted in increased tumor cell death. DFP also exhibits strength as a chelator with an ability to limit tumor growth, migration, and metabolism. DFP was found to inhibit overall cellular respiration capacity corresponding positively with the increase of dosage while generating ROS in the same pattern. DFX was shown to inhibit cell cycle progression, while downregulating proliferative pathways in gastric cancer cells.
In addition to DFO and its derivatives, other compounds with chelating abilities have gained attention with regard to cancer treatment. Shang et al. demonstrated that the chelators ciclopirox olamine and Di-2-pyridylketone-4,4-dimethyl-3-thiosemicarbazone (Dp44mT) could effectively permeate cancer cells and inhibit proliferative signaling through the mammalian target of rapamycin pathway. Dp44mT is a newer chelator that has also been studied against a number of cancer cell types in controlled experimental trials. Using Dp44mT against osteosarcoma cells in vitro and in vivo, Li et al showed that the chelator was able to inhibit cancer cell vitality and proliferation likely through caspase-dependent apoptosis. In vivo xenograft transplantation resulted in an average decrease in tumor mass of 62.22% in Dp44mT treated mice at the 30-day sacrifice point. Moreover, Krishan et al. found that Dp44mT was able to disrupt metabolic processes in order to drain ATP levels to the point of AMP-activated protein kinase activation, which eventually leads to autophagy/catabolism.
Two novel chelators, known as the super-polyphenols 6 and 10, were observed by Ohara et al. to have anticancer properties comparable to DFO and DFXas evidenced by induced apoptotic activity. Notably, these beneficial effects came without the associated additional toxicity of DFO and DFX. More studies using the novel super-polyphenol chelators are needed to reinforce the results from Ohara et al., but the study appears to be in line with established anticancer properties of chelation. Curcumin has also been reported to have iron-chelating properties and was used in a recent study to determine its effects on caspase-dependent apoptosis. Results indicated that curcumin was indeed effective at chelating iron and inducing apoptosis as evidenced by the expression of apoptosis markers caspase-3 and caspase-9. However, there were also protective actions taken by the cells in order to counter the effects of curcumin.
A potential drawback in chelation therapy may be the unabated attempts of cancer cells to restore iron homeostasis. Chen et al. reported that the usage of DFO against breast cancer cells resulted in increased expression of the iron import proteins DMT1 and TfR1, leading to an overall increase of intracellular iron concentrations. This phenomenon was only observed in aggressive triple-negative breast cancer (TNBC) cell lines, while the estrogen receptor+ non-aggressive cell lines did not experience the same phenomenon of iron accumulation. Activation of the IL-6/phosphatidylinositol-3 kinase (PI3K)/ protein kinase B (PKB, also known as Akt) pathway was observed in DFO treated cells from the aggressive TNBC groups in contrast to the estrogen receptor+ positive groups. The IL-6/PI3K/Akt pathway is an inflammatory pathway that promotes cancer cell survival.IL-6/PI3K/Akt pathway regulated iron uptake protein expression is suspected to be responsible for iron accumulation in TNBC cell lines; however, the underlying reason for the exclusivity to TNBC cells is not clear.
In a study by Liu et al., DFO promoted cell viability through a different mechanism, HIF-1α, which is likely to have arisen as a consequence of an iron deficient state. Although these studies were performed in vitro, they present possible concerns over the potential side effects of a more general treatment such as the sequestration of iron as opposed to more specific pathway targeting methods. The ability of cells to adapt to their environment poses particular challenges for anticancer treatments. A possible counter to this may be through the use of combined chelation and other therapies. In an in vivo study by Lang et al. involving administration of the chelator DFO and a HIF-1α inhibitor known as Lificiguat (YC1), anticancer efficacy was compared between pancreatic cancer cells treated with and without YC1. Compared to chelation treatment alone, use of DFO with YC1 was more effective in destroying cells and inhibiting their ability to overcome the lack of iron. Resistance to therapy is not exclusive to chelators. The common chemotherapy agent cisplatin is met with resistance that can actually be overcome with the help of chelators.In addition to countering HIF-mediated resistance, combining chelators with other conventional chemotherapeutics has gained attention for studies in both basic science and clinical investigation to counter other forms of resistance and amplifying the effects of other drugs.
Iron chelators DFO and DFX have been reported to synergize with the pyrimidine analog 5-Fluorouracil (5-FU), a classic chemotherapy drug that disrupts DNA synthesis, to diminish esophageal cancer growth in vitro and in vivo. DFP and Dp44mT can synergize with 5-FU to treat glioblastoma and breast cancer, respectively. DFO was reported to synergize with platinum-based chemotherapy drugs in different cancer types includingovarian cancer, neuroblastoma, cervical cancer. DFX, Triapine and Dp44mT were able to synergize with cisplatin to treat TNBC, advanced-stage solid tumor malignancies and cisplatin acquired resistant lung cancer, respectively.
Chelation has been shown to synergize with inhibitors of topoisomerases, which are involved in DNA strand opening during replication, as well as inhibitors of poly-ADP-ribose polymerase (PARP), which is involved in DNA repair. The chelator triapine was able to suppress the growth of BReast CAncer gene-wild type and PARP inhibitor-resistant ovarian cancer cells. DFP enhanced the anti-cancer efficacy of the alkylating agent temozolomide that damages DNA. Tury et al. demonstrated that DFX can work successfully in combination with the alkylating agent cyclophosphamide against TNBCs in mouse models. Iron chelation has also displayed synergistic activity with the intercalating agent doxorubicin in both leukemia and solid tumors.
Combining chelators with radiation therapy has been shown to amplify the anti-cancer effects of radiation therapy. The mechanism of action has been suggested to result from an inability to recover from radiation damage due to a lack of iron. In several studies Kunos and colleagues investigated and discussed clinical applications of triapine in combination with radiochemotherapy for the treatment of gynecological cancers, namely cervical and vaginal cancers. One study showed that triapine in addition to radiochemotherapy produced a progression-free survival rate at 18 months of 67% as opposed to 25% without triapine. Another study observed a 3-year disease free survival rate of 80 percent, and an overall survival rate of 82 percent in cervical cancer patients. Comparing patients who received triapine in addition to cisplatin-radiotherapy as opposed to the radiochemotherapy without triapine showed a 15 percent difference in a 3-year estimated survival rate (92 vs 77, respectively).
In the aforementioned cases, the chelator or the complementary drug appears to make up where the other comes up short.
HIF
HIFs exist to counteract the environmental changes in a cell that result in a perceived state of hypoxia (Figure 3). In addition to the previously discussed increases in cellular iron content mediated by HIF-2α [See Metabolic changes in iron-mediated carcinogenesis], their power over cellular transformation extends further, which makes them a valuable target. HIF-2α may play a role in immune system evasion by tumors through the upregulation of cyclooxygenase-2 (COX2), resulting in a downstream increased production of prostaglandin E2 (PGE2), a molecule with immunosuppressive activities in the COX2/microsomal prostaglandin E synthase-1/PGE2 pathway.As previously mentioned, activation of the HIF pathways is a crucial adaptation mechanism for the continued survival of tumor cells and may arise as an obstacle in chelation treatment if unaccounted for. Therefore, research into methods of inhibiting HIF activation constitute an important area in developing treatments for cancer.
A number of drugs are currently being studied for their ability to inhibit the HIF pathways. These drugs use a variety of mechanisms including transcriptional and translational inhibition, inhibition of quaternary structure formation, and promotion of degradation. A review of tested HIF inhibitors identifies and summarizes the observed effects of HIF-1α and/or HIF-2α inhibitors and details their mechanisms of action. One of the inhibitors from this review was YC-1, which was used in a chelation therapy study as a means of inhibiting HIF-1α activity and enhancing iron chelation. This was one of several mentioned inhibitors that targeted transcriptional activity. Several other inhibitors reviewed were reported to target molecular chaperones such as heat shock protein 90, a process that can negatively impact HIF-1α and HIF-2α stability. Usage of the HIF inhibitor TAT-ODD-procaspase 3 (TOP3)was also shown to yield success in murine models of pancreatic cancer when combined with gemcitabine or TS-1, extending survival rates from 0 to 25% after 100 and 50 days for TOP3-gemcitabine and TOP3-TS-1 combinations, respectively.
Ubiquitination (Ubiquitylation): “Ubiquitination, also known as ubiquitylation, is an enzymatic process that involves the bonding of an ubiquitin protein to a substrate protein. This has sometimes been referred to as the molecular “kiss of death” for a protein, as the substrate usually becomes inactivated and is tagged for degradation by the proteasome through the attachment of the ubiquitin molecule.”10
The search for other potential inhibitors of HIF and its effective pathway can open more doors for therapeutics as well. The gene Parkin, known for its role in Parkinson's Disease, has been reported to have tumor suppressing capabilities, and increases in Parkin expression in mice models resulted in degradation of HIF-1α and resultant inhibition of tumor progression, possibly through a ubiquitination-degradation mechanism. Delivery of a modified, soluble therapeutic form of the protein into Parkinson's Disease model rodents successfully protected against advancement of Parkinson's disease. This may suggest that the protein could have therapeutic potential if it can maintain function with modifications for drug delivery.
Summary and Conclusion
Given the complexity of cancer, it is beneficial to find patterns within it in order to both prevent and treat the disease. Patterns of iron dependence in cancers open the door to a field of therapeutics that aims to target cancer cells with a greater specificity in order to both maximize therapeutic efficacy and avoid harm to healthy cells. Furthermore, understanding iron's nature as a pro-oxidant and its potential role as a collaborator in carcinogenesis may help to provide easy, beneficial preventative measures against cancer. While there are a number of studies discussed in this review that illustrate a connection between iron and cancer, there has yet to be a definitive mechanism of action elucidated. It is possible that there may not be a single route towards disease when it comes to iron and cancer.
Another potential mechanism for synergism when compounding a treatment regime with doxycycline:
“Lactoferrin administration showed a significant HIF inhibitory effect in the retinal neuronal cells. Oral administration of lactoferrin or conditional Hif1a gene deletion significantly reduced CNV volume compared to controls. Conclusions Lactoferrin has a therapeutic effect in a laser CNV model by suppressing the retinal HIF activity.”11
“In this study we evaluated the physiological activity of this importin α/β-inhibitor (Ivermectin) in the hypoxia response pathway. Treatment with Ivermectin decreases binding activity of HIF-1α to the importin α/β-heterodimer. Moreover, HIF-1α nuclear localisation, nuclear HIF-1α protein levels, HIF-target gene expression, as well as HIF-transcriptional activity are reduced upon Ivermectin treatment.”12
Cellular iron metabolism has many individual components that make up a very delicate system that ensures vitality by providing the essential nutrients and, inadvertently, potential toxicity. Understanding these pathways has enabled scientific knowledge to advance to the point of identifying the extent of metabolic modification and dysregulation that occur in cancer and knowing how to use it. Targeting TfR1 can be useful as increasing levels of the receptor enhances cellular import of iron or other TfR1 ligand-conjugated anticancer drugs. Increased hepcidin levels resulting in greater intracellular iron concentrations is the basis for hepcidin based treatment that could inhibit or degrade hepcidin and relieve the iron accumulation. Studies on hepcidin inhibition show potential, but they also suggest that it may not be a standalone option. Sequestering iron by use of chelators is a more heavily tested method of targeting iron in general by depriving cancer cells of the iron that they need for their rapid growth. Cells may counteract the lack of iron through induction of HIFs. HIF inhibition may be an effective complement to iron chelation and other methods of therapeutics by severing a useful lifeline in iron and/or oxygen lacking cells, but further studies of treatment combinations are needed to better illustrate the effects of various methods on cancer cells. Regulated cell death by means of ferroptosis is a promising way of using iron against cancer cells. Unlike chelation and other treatments that decrease the level of iron in cells, ferroptosis needs iron in the cells to unleash its toxicity. Therapeutic methods that can induce this phenomenon may target the vital antioxidant defenses of the cell, which then gives way to the ROS-led destructive processes. A recent review has summarized the currently available pharmacological agents targeting iron metabolism. Keeping an eye on naturally occurring activation and inhibition mechanisms from transcription to post-translation can help shape and guide future strategies. In vivo testing and clinical trials will be necessary to generate a clearer image of the true potential of these treatments.
Doxycycline as a cancer therapeutic
To complete this review, four papers discuss the anticancer properties of doxycycline.
In 1998, Fife et al found that, in vitro, doxycycline can significantly inhibit the growth of prostate and breast cancer tumors by the inhibition of matrix metalloproteinases (MMPs) and induction of apoptosis:13
Abstract
Prostate cancer is the most common form of cancer in older men and the major cause of death from prostate cancer is metastatic disease. The matrix metalloproteinases (MMPs) play a significant role in the growth, invasion and metastasis of many tumors, including those of the prostate. We previously demonstrated that doxycycline, a synthetic tetracycline, inhibits MMPs and cell proliferation and induces apoptosis in several cancer cell lines. We also demonstrated that in an in vivo model of metastatic breast cancer in athymic mice doxycycline inhibits tumor size and regrowth after resection. In the present study, gelatinolytic activity in the human prostate cancer cell line, LNCaP, was suppressed and significant inhibition of cell growth occurred after exposure to 5 or 10 μg/ml of doxycycline, while cell growth was normal in untreated cells. Radioisotope incorporation into proteins was reduced by doxycycline. DNA fragmentation, consistent with apoptosis, was demonstrated in cells treated with doxycycline. These data suggest that doxycycline may have potential utility in the management of prostate cancer.
1. Introduction
Prostate cancer is the most common cancer in American men over the age of 65 years and is the second leading cause of cancer deaths in these individuals. Most deaths from prostate cancer, as from other malignancies, result from metastatic disease. Numerous mechanisms are involved in metastasis, including increased activity of the matrix metalloproteinase (MMP) family of enzymes. MMPs degrade the extracellular matrix surrounding cancer cells, allowing them to spread beyond the primary site and MMPs also enhance angiogenesis, or new blood vessel formation, a crucial phenomenon in metastasis. Another factor contributing to metastasis is the relatively unrestrained growth characteristic of most tumors. Many cancer cells exhibit a diminution in apoptosis, or programmed cell death, which is normally involved in the removal of damaged or senescent cells and which appears to be regulated in part by oncogenes and tumor suppressor genes and their protein products. This suppression of apoptosis probably plays a role in the unchecked growth and spread of tumors.
The tetracyclines (TCNs) are a group of antibiotics that have an inhibitory effect on MMP activity. The mechanism of this inhibition remains unclear, but it has been postulated to result from at least one of the following: (a) binding of calcium at the active site of the enzyme, which is critical to MMP function; (b) steric hindrance in the interaction of the enzyme with TCN; or (c) binding of TCN to the enzyme substrate. We recently have obtained evidence that the synthetic TCN, doxycycline, also selectively inhibits mRNA synthesis of types I and IV collagenase and stromelysin, thus suggesting yet another possible mechanism for its inhibitory action. We now present data indicating that doxycycline kills human prostate cancer cells in vitro and that at least some of the observed cell death is due to enhanced apoptosis.
Fig. 1. Growth curves of LNCaP cells cultured in the presence of no doxycycline or 5 or 10 μg/ml of doxycycline. Note essentially complete cell death at days 4 and 6 in cultures incubated with a dose of 10 μg/ml.
3.2. Protein synthesis
The radiolabeling studies demonstrated that doxycycline inhibited general cell protein synthesis by approximately two- to four-fold compared with controls prior to cell death (Fig. 2).
Fig. 2. [3H]Leucine incorporation into protein in cells cultured in the presence of no doxycycline or 5 or 10 μg/ml of doxycycline.
Gelatinolytic activity is a measure of the pro-matrix metalloproteinase (MMP) activity of cancer cells. It is usually higher in metastatic cancer cells than ordinary cell lines. Lower is better:
3.3. Gelatin zymography
We examined the gelatinolytic activity of the prostate cancer cells in the presence and absence of 10 μg/ml of doxycycline using gelatin zymography. We demonstrated a 50–75% reduction in gelatinolytic activity in conditioned medium from doxycycline-treated cells compared with untreated cells (Fig. 3).
Fig. 3. Electrophoresis of aliquots of medium from each culture condition in 9% SDS-polyacrylamide gel containing 1% gelatin. Clearing represents gelatinase activity, which is strikingly reduced in samples from cells incubated with 10 μg/ml of doxycycline (lane C). Lane A, MW markers; lane B, control sample (no doxycycline).
3.4. DNA laddering
The DNA gels demonstrated laddering of DNA most prominently in samples from cultures treated with 5 μg/ml of doxycycline for 2 days and 10 μg/ml of doxycycline for 2, 4 and 6 days in culture, indicating the occurrence of apoptosis, or programmed cell death (Fig. 4).
Fig. 4. Apoptosis demonstrated by DNA fragmentation in 1.2% agarose gel. Lane 1, DNA ladder; lane 2, DNA from control cells cultured for 2 days; lane 3, DNA from control cells cultured for 4 days; lane 4, DNA from control cells cultured for 6 days; lane 5, DNA from cells cultured for 2 days with 5 μg/ml of doxycycline; lane 6, DNA from cells cultured for 4 days with 5 μg/ml of doxycycline; lane 7, DNA from cells cultured for 6 days with 5 μg/ml of doxycycline; lane 8, DNA from cells cultured for 2 days with 10 μg/ml of doxycycline; lane 9, DNA from cells cultured for 4 days with 10 μg/ml of doxycycline; lane 10, DNA from cells cultured for 6 days with 10 μg/ml of doxycycline. Note laddering in lanes 5, 8, 9 and 10, consistent with fragmentation of DNA.
4. Discussion
We have been studying the effects of doxycycline on several types of cancer cells, specifically human breast cancer cells (MDA-MB-435), human prostate adenocarcinoma cells (LNCaP) and human osteosarcoma cells (U20S), and have found inhibition of cell proliferation, gelatinolytic (MMP) activity, MMP mRNA synthesis and Matrigel invasion (an in vitro surrogate for metastasis). We previously conducted a trial of doxycycline administered either before implantation of breast cancer cells into athymic mice or at the time of resection of the primary tumors and showed significant reduction in the size of the primary tumors in the pretreated animals and in the number of lung metastases in all animals.
In the present study, we have demonstrated that doxycycline kills prostate cancer cells and that at least some of this cell death is due to apoptosis, which is demonstrated by DNA laddering. Apoptosis is a normal mechanism for eliminating senescent or diseased cells and is suppressed in many cancers by various factors, including oncogene and tumor suppressor proteins. The ability to `turn on' apoptosis in cancer cells has great potential for inducing tumor remission. It is unclear at present whether there is any relationship between the effects of doxycycline on MMP activity and the induction of apoptosis; indeed, it is likely that these represent two discrete aspects of doxycycline activity. We also have shown elsewhere that doxycycline induces apoptosis in vitro in human osteosarcoma cells and in human breast cancer cells.
The potential ramifications of these data are significant. If a relatively safe well-tolerated agent such as doxycycline can inhibit cell proliferation and induce apoptosis, it could represent a major addition to the treatment options available for patients with prostate cancer, especially for adjuvant therapy, and could reduce the metastatic burden in animals and, ultimately, in humans. We have begun a phase I clinical trial in humans to determine the best tolerated dose of doxycycline in patients with prostate cancer. Preliminary results indicate that serum levels of 5–10 μg/ml can be safely achieved with doses that are tolerated by most patients.
No conflict of interest statement was provided.
From 2016, Zhang et al conducted an in vitro study using human breast cancer cell lines:14
ABSTRACT
Experimental evidence suggest that breast tumors originate from breast cancer stem cells (BCSCs), and that mitochondrial biogenesis is essential for the anchorage-independent clonal expansion and survival of CSCs, thus rendering mitochondria a significant target for novel treatment approaches. One of the recognized side effects of the FDA-approved drug, doxycycline is the inhibition of mitochondrial biogenesis. Here we investigate the mechanism by which doxycycline exerts its inhibitory effects on the properties of breast cancer cells and BCSCs, such as mammosphere forming efficiency, invasion, migration, apoptosis, the expression of stem cell markers and epithelial-to-mesenchymal transition (EMT) related markers of breast cancer cells. In addition, we explored whether autophagy plays a role in the inhibitory effect of doxycycline on breast cancer cells. We find that doxycyline can inhibit the viability and proliferation of breast cancer cells and BCSCs, decrease mammosphere forming efficiency, migration and invasion, and EMT of breast cancer cells. Expression of stem cell factors Oct4, Sox2, Nanog and CD44 were also significantly downregulated after doxycycline treatment. Moreover, doxycycline could down-regulate the expression of the autophagy marker LC-3BI and LC-3BII, suggesting that inhibiting autophagy may be responsible in part for the observed effects on proliferation, EMT and stem cell markers. The potent inhibition of EMT and cancer stem-like characteristics in breast cancer cells by doxycycline treatment suggests that this drug can be repurposed as an anti-cancer drug in the treatment of breast cancer patients in the clinic.
KEYWORDS: autophagy, breast cancer, cancer stem cells, doxycycline, epithelial-to-mesenchymal transition, mitochondria
Inhibition of mitochondrial biogenesis is one of the mechanisms:
Doxycycline is a tetracycline-derivative wide-spectrum antibiotic, which was first approved by FDA in the late 1960s. Tetracyclines inhibit protein synthesis by interfering with the binding of activated aminoacyl-tRNAs on the A-site of the 30S subunit of bacterial ribosomes. The 30S bacterial ribosome is homologous to the 28S mitochondrial ribosome in mammalian cells, leading to manageable side effects from tetracycline treatment, by inhibiting mitochondrial biogenesis in mammalian cells. Interestingly, recent studies have demonstrated that BCSCs rely more on oxidative phosphorylation. We found that BCSCs produce less lactate and have higher ATP content and mitochondrial labeling compared to their differentiated progeny. Supporting a dependency of BCSCs on mitochondrial oxidative phosphorylation, more recently, Lamb et al. found that mammospheres (cell cultures enriched for BCSCs) highly upregulate mitochondrial proteins, and that doxycycline can inhibit mitochondrial biogenesis and the “stem cell” phenotype of breast cancer cells.
Targeted cancer and stem cell effects, but they weren’t exactly sure why and chelation hasn’t even been referred to:
However, although Lamb et al. demonstrated an effect of doxycycline on the CSC phenotype of a panel of different cancer cell lines (including 2 breast cancer lines), these observations were limited to only tumor-sphere forming assays. Therefore, the mechanism by which doxycycline exerts its inhibitory effects on the properties of breast cancer cells and BCSCs remain unclear. We build on the previous studies and in the present study investigated the inhibitory effect of doxycycline on the growth, migration, invasion, epithelial-to-mesenchymal transition (EMT), expression of stem-cell factors and autophagy in breast cancer cells in vitro.
Doxycycline inhibits cell viability of breast cancer cells. (A) Breast cancer cells propagated as monolayers (differentiated cultures, solid line) or mammospheres (enriched in BCSCs, dotted line) were treated with the indicated concentrations of doxycycline for 72h. The % viability of cells at each doxycycline dose at the end of treatment was measured using a Cell Counting Assay Kit. The horizontal dotted line represents the value of IC50 (MCF7 and MDA-MB-468 was 11.39 and 7.13uM, respectively). (B) Doxycycline significantly reduced the number of colonies in both breast cancer cell lines, MCF7 and MDA-MB-468. (C-D) Doxycycline treatment induced both early (Annexin-Vpos/PIneg) and late (Annexin-Vpos/PIpos) apoptotic cell death in both breast cancer lines.
Doxycycline inhibits self-renewal of breast cancer cells. Doxycycline decreased the % of CD44+CD24-/low BCSC population in MCF7 and MDA-MB-468 cells (A), as well as mammosphere formation efficiency in both lines tested (B). (C) Relative mRNA expression levels of self-renewal related genes are significantly downregulated in doxycycline treated breast cancer cells. The dotted line represents mRNA expression of non-treated breast cancer cells. (D) Western blot analysis of the stem cell related genes. (Data are reported as means ± standard deviation, *p < 0.05, **p < 0.01) MCF7 and MDA-MB-468 were treated with 11.39 μM and 7.13 μM doxycycline, respectively.
Doxycycline inhibits invasion, migration, and epithelial-to-mesenchymal transition of breast cancer cells
BCSCs have been shown to have an invading phenotype therefore, next we investigated whether the inhibition of viability by doxycycline treatment affected the invasion and migration capabilities of breast cancer cells. We performed transwell invasion and migration assays in the absence and presence of matrigel basement membrane. MCF7 cells have relatively low migration and invasion abilities therefore, we choose the MDA-MB-468 for these studies. Results showed that a 72-hour pre-treatment with doxycycline significantly inhibits their invading and migrating abilities (Fig. 3). Migration and invasion efficiencies were reduced by 52.08% (p = 0.023) and 52.88% (p = 0.0043, Students paired, 2-tailed t-test) respectively.
Doxycycline inhibits migration and invasion in MDA-MB-468 breast cancer cells. Doxycycline inhibits migration (A) and invasion (B) in MDA-MB-468 breast cancer cells in a transwell assay. MDA-MB-468 cells were treated with doxycycline for 72 h with a single dose of IC50.
Doxycycline decreased the EMT phenotype in MDA-MB-468 breast cancer cells. (A) Relative mRNA expression levels of EMT-related genes in doxycycline treated breast cancer cells. The dotted line represents mRNA expression of non-treated breast cancer cells. (Data are reported as means ± standard deviation, *p < 0.05, **p < 0.01) (B) Western-blot analysis for EMT-related proteins. MDA-MB-468 cells were treated with doxycycline for 72 h with a single dose of IC50.
Doxycycline suppresses autophagy markers
Autophagy has been shown to suppress tumor initiation at an early stage however, it can also help cancer cells survive under hypoxia, under-nutrition, antitumor therapies, and other stress conditions and is considered a general feature of solid tumors. Earlier reports have also demonstrated an important role for autophagy in the maintenance of CSCs and metastasis. Thus, we decided to analyze the effect of doxycycline on 2 autophagy-related proteins, LC-3BI and LC-3BII, as 2 of the most specific biomarkers of autophagy with broad tissue specificities and widely used in autophagy-related studies. Treatment with a single dose of doxycycline resulted in suppression of protein levels of LC-3BI and LC-3BII in both cell lines tested (Fig. 5A-B, Students unpaired, 2-tailed t-test), suggesting a potential mechanism by which doxycycline treatment mediates suppression of self-renewal in breast cancer stem cells.
Doxycycline inhibits decreases autophagy-related protein levels. LC3BI and LC3BII protein levels were analyzed (A) and measured (B) in MCF-7 and MDA-MB-468 cells after doxycycline treatment. MCF7 and MDA-MB-468 were treated with 11.39 and 7.13 μM doxycycline for 72 h, respectively.
Discussion
An increasing body of evidence demonstrates that breast cancer cell populations enriched for cells that express “stem cell” markers have significantly higher tumor-forming capacity, and we have recently shown that this subpopulation of breast cancer cells is important not only for tumor initiation, but also propagation. It is now believed that elimination of BCSCs is necessary to achieve long-term tumor control.
These findings have launched an effort for identifying the Achilles heel of CSCs with the goal of developing anti-cancer drugs that not only eliminate the more differentiated cells within tumors, but also effective against the CSC population. Recently, Lamb et al. used an unbiased quantitative proteomic profiling to identify the global phenotypic properties of cancer stem cells (CSCs) that could be targeted across multiple tumor types. They found that mitochondrial biogenesis was essential for the anchorage-independent clonal expansion and survival of CSCs, so this common feature could be utilized to target CSCs and treat cancer effectively as a single disease of “stemness”.
In particular, doxycycline significantly inhibits proliferation and BCSC self-renewal ability in MCF7 and MDA-MB-468 cell lines, with IC50 values similar to those reported by Lamb et al.7 Of note, and in agreement with previous findings serum-free mammosphere cultures enriched in BCSCs were more resistant to doxycycline compared to the more differentiated monolayer cultures propagated in serum-supplemented media (Fig. 1A). Furthermore, we demonstrated that this old drug was very efficient at eliminating the CD44+CD24−/lowBCSC population as shown in Figure 2A.The observation that doxycycline can eliminate the BCSC population identified by the surface marker profile (CD44+CD24−/low) was confirmed by functional mammophere formation assays (Fig. 2B). Finally, the inhibition of self-renewal of BCSCs by doxycycline was accompanied by a significant downregulation of stem cell-associated factors, such as Oct4, Sox2, Nanog, c-myc and CD44 at the gene transcriptional level (Fig. 2C) as well as protein level (Fig. 2D). It should be emphasized, that since CD44+CD24−/lowBCSCs display a mesenchymal state with high invasive capacity, our results on the effect of doxycycline on BCSCs mainly apply to the mesenchymal-like BCSCs, and that these conclusions do not necessarily apply to epithelial-like, ALDH-pos BCSCs.
BCSC and EMT contributes to metastasis in breast cancer. EMT refers to the initial stage of invasion and metastasis, during the process of metastasis, during which the polarized epithelial cells convert into motile mesenchymal cells. The decrease expression of E-cadherin, which often occurs concurrently with increased expression of N-cadherin, is a critical step in the EMT process. Expression of vimentin represents the completely dedifferentiated state of tumor cells. Recently, Qin Y et al. reported that doxycycline could reverse EMT and suppress the proliferation and metastasis of lung cancer cells.Wan L et al. reported that combination of aspirin, lysine, mifepristone and doxycycline have an inhibitory effect on the expression of cell adhesion molecules and cancer metastasis. In the present study, we found that doxycycline treated breast cancer cells have higher expression levels of E-cadherin, accompanied with lower expression levels of N-cadherin and vimentin compared to the nontreated cells, indicating a reversal of EMT (Fig. 3 and 4). These molecular changes were also associated by changes in migration and invasion ability, which were significantly inhibited by a single treatment with doxycycline (Fig. 3).
Repositioning of old drugs to new indications is an attractive approach as it has the potential to save considerable effort and time involved in the drug development process. Doxycycline is an FDA-approved antibiotic which has been used to treat a wide variety of bacterial and parasitic infections for nearly 50 years, and importantly without significant side effects. It has been found that doxycycline can down-regulate DNA-PK and radiosensitize tumor initiating cells and Pulvino et al. have reported that doxycycline could inhibit tumor growth of diffuse large B-cell lymphoma. In combination with the study presented here, these studies make doxycycline an attractive candidate for repurposing for cancer treatment. In the clinic, doxycycline has favorable pharmacokinetics with a long serum half-life (18-22h), when a standard dose of 200mg per day is administered.
In summary, our findings support that doxycycline, a FDA approved drug, can inhibit proliferation of breast cancer cells, as well as self-renewal of BCSCs. Part of the mechanism of inhibition of self-renewal is due to the suppression of key developmental stem cell factors, such as Oct4, Sox2, Nanog and c-myc. Importantly, treatment with doxycycline results in inhibition of EMT, invasion and migration of breast cancer cells, as EMT-related markers are significantly down-regulated. Finally, doxycycline treatment suppresses autophagy-related proteins, suggesting a role autophagy in the doxycycline-induced effects. In light of this study it is plausible to envisage clinical treatment strategies for breast cancer patients that include this old drug to prevent relapse and metastasis, and improve prognosis of breast cancer patients.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
This work was supported by China council scholarship.
This paper by Zhu et al (2017) conducted an in vitro investigation into how doxycycline synergizes with the chemotherapeutic drug doxorubicin to inhibit the proliferation of castration-resistant prostate cancer cells, a condition that was previously untreatable:15
Abstract
Castration-resistant prostate cancer (CRPC) is fatal and there is currently no effective clinical treatment. The antibiotic doxycycline has shown anti-cancer effect in several kinds of solid tumors including prostate cancer. In this study, a combination of doxycycline and doxorubicin was used to investigate the synergistic effect on CRPC cells. MTT assay was employed to determine the viability of cells in two-dimensional (2D) cultures. Apoptosis was determined by Annexin V/propidium iodide (PI) double staining assay. Cell cycle was analyzed by PI staining, and reverse transcription-PCR (RT-PCR) was used to determine the expressions of apoptosis-related genes at mRNA level. Western blot analysis was used to analyze the expressions of Bcl-2, Bax, and Poly (ADP-ribose) polymerase proteins. Cytotoxicity assay and morphological observation of PC3 cells in three-dimensional (3D) cultures were used to determine the effect of combination treatment. Results showed that doxycycline combined with doxorubicin significantly inhibited PC3 cells in both 2D and 3D cultures, enhanced apoptosis, and increased the accumulation of cells in G2/M phase. RT-PCR showed down-regulation of Bcl-2 and up-regulation of Bax mRNA after combination treatment. Meanwhile, western blot analysis showed that combination treatment resulted in down-regulation of Bcl-2 protein and up-regulation of Bax protein, and that PARP cleavage was obviously exhibited after combination treatment. Confocal imaging analysis indicated that doxorubicin penetrated deeply into the core of spheroids when combined with doxycycline. These data indicated that doxycycline in combination with doxorubicin had a synergistic effect on PC3 cells and may provide a potential novel strategy for the treatment of CRPC.
Efficacy against different castration resistant prostate cancer cell lines. Concentration dependent action. Higher % inhibition is better. C, (LNCaP) is a non-CRPC cell line:
The synergistic effects of DC and DOX on the PC3, DU145, and LNCaP cells (A) PC3 cells treated with a combination of DC (1.25 μM) and DOX (40–320 nM). (B) DU145 cells treated with a combination of DC (1.25 μM) and DOX (40 to 320 nM). (C) LNCaP cells treated with a combination of DC (1.25 μM) and DOX (40–320 nM). (D) The effects of combination treatment with DC (1.25 μM) and DOX (80 nM) on PC3, DU145, LNCaP, and LO2 cells. **P < 0.01, *P < 0.05, ‘ns’: no significance vs. control.
Meanwhile, MTT assay was also used to detect the effect of combination treatment on human normal hepatocytes LO2 cells. As shown in Fig. 1D, the combination of DC (1.25 μM) and DOX (80 nM) resulted in the lowest inhibition rate in the LO2 cells, compared with that in the PC3, DU145, and LNCaP cells, respectively, indicating that the combination treatment of DC and DOX had low cytotoxicity to normal cells at the concentrations of drugs used in the study.
Higher % apoptosis is better:
Effects of combination treatment on the apoptosis and cell cycle arrest in the PC3 cells PC3 cells were treated with the combination of DC (1.25 μM) and DOX (80 nM). (A) Cell apoptosis was analyzed by flow cytometry. (B) Quantitative analysis on the apoptosis of cells. (C) Analysis of cell cycle distribution. **P < 0.01, *P < 0.05 vs. control.
These results indicated that combination treatment significantly increased the number of PC3 cells arrested in the G2/M phase of cell cycle, compared with single-drug treatment.
Effects of combination treatment of DC and DOX on the mRNA expressions of apoptosis-related genes (A) The combination treatment of DC (1.25 μM) and DOX (80 nM) induced the up-regulation of Bax and down-regulation of Bcl-2 at the mRNA level, and GAPDH was used to normalize the expression of genes. (B) Quantitative analysis of Bcl-2 mRNA expression level. (C) Quantitative analysis of Bax mRNA expression level. **P < 0.01, *P < 0.05 vs. control.
The results showed that the combination of DC and DOX significantly decreased the expression of Bcl-2 mRNA (45.2% ± 1.6%), compared with that in the single-DC (87.2% ± 3.3%, P < 0.01) and single-DOX (76.1% ± 2.1%, P < 0.01) groups, respectively. Meanwhile, the expression level of Bax mRNA (180.2% ± 2.8%) was significantly increased in the combination group, compared with that in the single-DC (127.5% ± 3.3%, P < 0.01) and single-DOX (154.0% ± 2.6%, P < 0.01) groups, respectively, indicating that the combination treatment of DC and DOX synergistically regulated the expression of apoptosis-related genes at mRNA level.
“Bcl-2 (B-cell lymphoma 2), encoded in humans by the BCL2 gene, is the founding member of the Bcl-2 family of regulator proteins that regulate cell death (apoptosis), by either inhibiting (anti-apoptotic) or inducing (pro-apoptotic) apoptosis. It was the first apoptosis regulator identified in any organism.”
“Apoptosis regulator BAX, also known as bcl-2-like protein 4, is a protein that in humans is encoded by the BAX gene. BAX is a member of the Bcl-2 gene family. BCL2 family members form hetero- or homodimers and act as anti- or pro-apoptotic regulators that are involved in a wide variety of cellular activities. This protein forms a heterodimer with BCL2, and functions as an apoptotic activator. This protein is reported to interact with, and increase the opening of, the mitochondrial voltage-dependent anion channel (VDAC), which leads to the loss in membrane potential and the release of cytochrome c. The expression of this gene is regulated by the tumor suppressor P53 and has been shown to be involved in P53-mediated apoptosis.”
Effects of combination treatment of DC and DOX on the expressions of Bcl-2, Bax, and PARP proteins (A) The combination treatment of DC (1.25 μM) and DOX (80 nM) resulted in the up-regulation of Bax and cleavage of PARP proteins and down-regulation of Bcl-2 protein. β-Actin was used to normalize the protein expression. (B) Quantitative analysis of Bcl-2 proteins. (C) Quantitative analysis of Bax proteins. (D) Quantitative analysis of PARP proteins. **P < 0.01, *P < 0.05 vs. control.
Combination treatment regulated the expressions of Bcl-2, Bax, and PARP proteins in PC3 cells
To further elucidate the mechanism underlying the effect of combination treatment on PC3 cells, western blot analysis was used to determine the expressions of Bcl-2, Bax, and PARP proteins after combination treatment. The results were analyzed by Image J software. As shown in Fig. 4A–C, combination treatment significantly down-regulated the expression of Bcl-2 protein (70.7% ± 3.3%) and up-regulated that of Bax protein (135.7% ± 3.7%), compared with the results in the single-DC (84.0% ± 3.2%, P < 0.05 and 115.7% ± 2.9%, P < 0.05, respectively) and single-DOX group (81.7% ± 2.6%, P < 0.05 and 111.0% ± 1.4%, P < 0.01, respectively). Meanwhile, Fig. 4D showed that PARP cleavage was also obviously enhanced in the combination treatment group (433.6% ± 7.6%), compared with that in the single-DC (206.6% ± 7.9%, P < 0.05) and single-DOX (201.5% ± 2.5%, P < 0.01) groups, respectively, suggesting that combination treatment significantly induced the apoptosis of the PC3 cells.
Combination treatment was effective on spheroids in 3D cell cultures
As shown in Fig. 5, in 3D cell cultures, DOX alone had no effect on the morphology, and only slight cytotoxic activity against spheroids (4.33% ± 0.56%, P < 0.01) was observed after treatment with DOX alone. However, DC alone made the spheroids disassociate into loose clusters of cells and showed higher cytotoxicity against spheroids (17.13% ± 4.06%, P < 0.05). Moreover, compared with DOX alone or DC alone, the combination treatment showed the most significant effect on disassociation of the spheroids. Furthermore, cytotoxic activity was enhanced when the spheroids were subject to combination treatment (30.67% ± 4.16%). These results indicated that combination treatment had greater cytotoxic activity and was more effective in inducing disassociation of the spheroids.
Effects of combination treatment of DC and DOX on the morphology and cytotoxicity against spheroids formed by PC3 in 3D cultures (A) The morphology of spheroids treated with DC alone (1.25 μM), DOX alone (80 nM), or a combination of DC (1.25 μM) and DOX (80 nM) for 6 days in 3D cultures. The spheroids without drug treatment were used as the control. Scale bars = 200 μm. (B) Spheroids were treated with DC alone (1.25 μM), DOX alone (80 nM), or a combination of DC (1.25 μM) and DOX (80 nM) for 6 days in 3D cultures. **P < 0.01, *P < 0.05 vs. control.
The spheroids are constituted by organized extracellular matrix such as fibronectin, laminin, and collagen. And the spheroids formed in 3D cultures possess characteristics of tumor more similar to in vivo. As a model to test the effectiveness of chemotherapy, 3D culture assay was used to investigate the efficacy of drug delivery system and distribution of drug in the spheroids. Here, 3D cultures were used to further explore the efficacy and underlying mechanism of combination treatment on the PC3 cells cultured in 3D environment. In 3D cultures, low-dose DOX alone had no effect on the morphology of spheroids, and was only slightly effective in inducing cytotoxic effects after single-DOX treatment. The reason for the insensitivity of spheroids to DOX alone may be that cancer cells cultured in the 3D environment are more resistant to various anti-cancer drugs. However, obvious disassociation of the spheroids and enhanced cytotoxic activity were observed in the single-DC group, compared with that of in single-DOX group, indicating that in addition to its cytotoxic effectiveness, DC alone could disrupt the organization of spheroids.
Conclusion
In summary, we have revealed that DC enhances the effectiveness of DOX against CRPC cells both in 2D and 3D cultures, and the combined treatment of DC and DOX may provide a new potential therapeutic strategy for future clinical treatment.
Our final paper. From 2019, Markowska et al conducted a review into the repositioning of doxycycline, salinomycin, monensin and ivermectin as cancer drugs.16
I will focus on their research into doxycycline and ivermectin, in line with current treatment protocols being used in an ongoing large scale community trial:
Chemotherapy is one of the standard methods for the treatment of malignant tumors. It aims to cause lethal damage to cellular structures, mainly DNA. Noteworthy, in recent years discoveries of novel anticancer agents from well-known antibiotics have opened up new treatment pathways for several cancer diseases. The aim of this review article is to describe new applications for the following antibiotics: doxycycline (DOX), salinomycin (SAL), monensin (MON) and ivermectin (IVR) as they are known to show anti-tumor activity, but have not yet been introduced into standard oncological therapy. To date, these agents have been used for the treatment of a broad-spectrum of bacterial and parasitic infectious diseases and are widely available, which is why they were selected. The data presented here clearly show that the antibiotics mentioned above should be recognised in the near future as novel agents able to eradicate cancer cells and cancer stem cells (CSCs) across several cancer types.
Keywords: Anticancer antibiotic; Cancer stem cells; New anticancer agent; Off-target effect.
Introduction
…Over the last several years, antibiotics with cytostatic activity have been registered as antineoplastic therapy agents. The most commonly applied include: doxorubicin, actinomycin, mitoxantron, bleomycin and mitomycin. DNA is the most common molecular target. These drugs affect DNA synthesis and replication through interference in a DNA sequence, interaction with DNA by intercalation and through the inhibition of topoisomerase all of which prevent the cancer cells from further division.
Recently, novel anticancer agents have been identified among numerous antibiotics, including those selected for this study. We are of the opinion that such discoveries can pave the way for new appealing treatments for neoplastic disease. Therefore, in this short review article we discuss the possibility of the off-label use of doxycycline (DOX), salinomycin (SAL), monensin (MON) and ivermectin (IVR), which are currently used for the treatment of a broad-spectrum of bacterial and parasitic infectious diseases.In vitro and in vivo studies of the anticancer activity of these drugs suggest promising clinical implications for their use as new anticancer agents.
Doxycycline
Doxycycline (DOX, Fig. 1) introduced in 1967, is a semisynthetic derivative of oxytetracycline. There are, however, many differences between DOX and natural tetracyclines, including markedly different pharmacokinetic properties, resulting in both lower doses and less frequent administration. DOX shows a broad spectrum of antibacterial activity. It is well tolerated, almost 100% is absorbed in the alimentary tract and it effectively penetrates the blood-brain barrier. The major target of antibiotics such as tetracyclines is the ribosome and protein synthesis within the cell resulting in the prevention of the binding of aminoacyl t-RNA to the 30S ribosomal subunit.
Fig. 1. The chemical formula of DOX.
DOX has a broad spectrum of antimicrobial activity that includes many respiratory pathogens and an apparent excellent safety profile. It was initially widely used to treat young children with respiratory tract infections. Moreover, DOX has a good safety record when used for a long time and is well-tolerated in humans. DOX is also safe to use in pregnancy and early childhood because no correlation was noted between its use and teratogenic effects during pregnancy.
In addition to its antibacterial activity DOX has cytotoxic and anti-proliferating properties in various cancer cells. Current studies have demonstrated that DOX is also a pluripotent drug that affects a wide range of pleiotropic therapeutic properties such as the control of invasive and metastatic cancer cells including anti-tumor growth effect and the inhibition of migration of cancer cells. It also exhibits interesting potential regarding enhanced therapeutic activity of several cancer therapies. DOX inhibits the growth of tumor cells and metastases. Using established cervical cancer cell lines (HeLa), Yang et al.20 documented that DOX inhibits proliferation, induces apoptosis and reduces invasion of tumor cells. It also decreases cancer stem cell markers: SOC-2, OCT-4, NANOG, NOTCH and BMI-1 in cell culture. In an animal model DOX reduces the cell proliferation markers: Ki67 and PCNA. This observation is significant as cancer stem cells (CSCs) are linked to resistance to treatment with cytometastatics, thought to be responsible for the relapse of the disease.
The antineoplastic effect of DOX, synergistic with the commonly applied anticancer drug doxorubicin, is also noted in prostate cancer cells: cancer cells underwent apoptosis due to an increase in the antiapoptotic Bax protein and a reduction in the antiapoptotic Bcl-2 protein. A similar mechanism of DOX action was noted in an animal model of pancreatic cancer: a reduction in the antiapoptotic protein as well as lowered expression of angiogenic IL-8.
Another mechanism involves the inhibition of leukemic cell migration due to a lower expression of matrix metalloproteinase MMP2 and MMP9 associated with cellular migration and the inhibition of adhesive phosphorylation FAK (focal adhesion kinase). Lee et al.5 studied the effect of DOX on cell lines of fibroneuroma that developed on the basis of neurofibromatosis (NF-1) (malignant peripheral sheath tumor). DOX was additionally applied alongside photodynamic therapy (PDT) and 5-aminolevulinic acid (ALA) in the treatment of this disease. DOX was associated with reduced toxic symptoms that occur in photodynamic therapy. The authors recommend further clinical studies with oral DOX during local PDT.
Ivermectin
Ivermectin (IVR, Fig. 4) is a 22,23-dihydro derivative of avermectin B1 (Fig. 4) from macrocyclic lactone produced by the Streptomyces avermitilis bacterium. IVR, recognised in the 2015 Nobel Prize in Physiology or Medicine, is a strong antiparasitic agent and almost four decades after its remarkable commercial introduction in 1981 for the control of endoparasitic nematodes and ectoparasitic arthropods in livestock, IVR was FDA-approved for human use in 1987.
Fig. 4. The chemical structure of avermectins and IVR. The two component chemicals are extremely similar, ivermectin B1a has an ethyl group at the C-26 position, while ivermectin B1b has a methyl group. IVR is composed of at least 80% of B1a and not more than 20% of B1b.
IVR consists of a mixture of two homologues containing at least 80% 5-O-demethyl-22,23-dihydroavermectin and less than 20% 5-O-demethyl-25-de(1-methylpropyl)-22,23-dihydro-25-(1-methylethyl)avermectin, generally referred to as 22,23-dihydroavermectin B1a and B1b, respectively (Fig. 4), and it is obtained through selective, catalytic hydrogenation of the cis-22,23-double bond of the avermectins B1a and B1b.
IVR belongs to the group of broad-spectrum antiparasitic agents which have a unique mode of action and is currently authorized to use for the treatment of onchocerciasis, lymphatic filariasis, strongyloidiasis, scabies and head lice. Recently, it has been shown that IVR can also exhibits a lot of new interesting activities such as antibacterial, antiviral and anticancer. IVR acts as a positive allosteric regulator of several channels including the glutamate-gated chloride channel (GluCl), γ-aminobutyric acid type-A receptor, glycine receptor, neuronal α7-nicotinic receptor and purinergic P2X4 receptor. In most of the IVR-sensitive channels, the effects of IVR include the potentiation of agonist-induced currents at low concentrations and channel opening at higher concentrations. IVR also acts as a positive allosteric regulator of several ligand-gated ion channels in vertebrates. Submicromolar concentrations of IVR activate or modulate the γ-aminobutyric acid type-A receptor (GABAAR), glycine receptor (GlyR) and neuronal α7-nicotinic receptor (nAChR). Its anti-parasitic activity is strictly connected with selective binding and high affinity of this compound to the GluCl channels which occur in invertebrate nerve and muscle cells. This leads to an increase in the permeability of the cell membrane to chloride ions with hyperpolarization of the nerve or muscle cell, resulting in paralysis and death of the parasite. The basis of IVR activity is the fact that some mammals do not have GluCl channels, that IVR has a very low affinity to mammalian GluCl channels and that it does not cross the blood-brain barrier in humans. IVRappears safe for human use, though there have been reports describing parasympathetic disturbances linked to the drug (salivation, dilation of pupils). In addition to well-known anti-parasitic activity of IVR, this compound has been recently shown to exhibit potent anti-cancer activities and may have substantial value for the treatment of a variety of cancers. IVR manifests antineoplastic activity related to its ability to inhibit multidrug resistance (MDR) proteins, the AKT/mTOR pathway and blocking the Wnt/TCF pathway (transcription factor of T-cells). IVR causes the degradation of PAK-1 (p21 – activated kinase), a main oncogenic kinase. It is effective in colon cancer, glioma multiforme and melanoma as well as skin and lung cancer by Wnt-TCF blocking.
IVRalso increases the level of intracellular ROS (reactive oxygen species) in tumor cells, associated with oxidative stress and DNA damage. Additionally, it preferentially inhibits cells resembling breast cancer stem cells (CSCs). Exposure to IVR reduces the expression of markers typical of stem cells: NANOG, OCT-4 and SOX2 (also the transcription factor of stem cells on the levels of mRNA and protein). This action is similar to that of SAL described earlier.
Dou et al. have shown that IVR induces autophagy, a self-degrading effect in breast cancer. Studies conducted on cell lines of breast cancer and on animal models, plus breast cancers of 20 patients have demonstrated reduced autophagy of breast cancer cells linked to reduced expression of PAK-1 due to the ubiquitin mediated degradation. The inhibition of PAK-1 reduced the phosphorylation of Akt, leading to the Akt/mTOR signalling pathway blocking, with the resulting decrease in tumor growth.
In other studies, IVR has been demonstrated to selectively inhibit SIN 3 – a protein that is associated with the pathogenesis of triple negative breast cancer. Using stabilized cell lines of ovarian cancer, Hashimoto et al. have proved that IVR induces the inactivation of PAK-1 kinase, intensely inhibiting the growth of ovarian cancer. The same authors have also observed an inhibition of malignant neuroma growth with no effect on the normal cell line. IVR was found to inhibit the YAP1 protein (yes-associated protein 1), whose the nuclear accumulation is linked to poor prognosis in gastric cancer. Using in vitro proliferation and animal model tests, IVR was shown to manifest a promising therapeutic potential in the inhibition of gastric cancer due to blocking of YAP1.
Conclusions
Anticancer antibiotics have made an important contribution to the area of antitumor chemotherapeutics. Different classes of antibiotics such as anthracyclines (daunorubicin, doxorubicin, epirubicin), glycopeptides (bleomycins), indolocarbazoles (staurosporine), exhibit anti-cancer properties. The antibiotics which are currently used in anticancer chemotherapy act in various ways, influencing molecules and signalling pathways.
The last decade has seen increased awareness of known drugs including antibiotics repositioned as antineoplastic agents and the results of further clinical trials regarding their efficacy are awaited. Research in this field should not only focus on developing newer safer derivatives of known derivatives of doxycycline (DOX), salinomycin (SAL), monensin (MON) and ivermectin (IVR) but also investigate novel drug delivery systems (NDDS). The evidence presented in this short review article indicates that the discussed compounds (DOX, SAL, MON, IVR) could be promising scaffolds in the development of new therapeutic strategies in cancer therapy in the near future. Significant changes in the parent chemical structures of these compounds can also lead to progress in this field. Future clinical trials for testing the efficacy of these cancer-targeted antibiotics, in multiple cancer types, are now clearly clinically warranted. The use of antibiotics in anti-cancer therapy can also be cost-effective therefore making treatment more accessible in the developing world.
Dosing and contraindications
Doxycycline Monohydrate - Uses, Side Effects, and More17
COMMON BRAND(S): ACTICLATE, MONODOX, VIBRA-TABS, VIBRAMYCIN
GENERIC NAME(S): DOXYCYCLINE MONOHYDRATE
How to use doxycycline monohydrate oral
Read the Patient Information Leaflet if available from your pharmacist before you start taking doxycycline and each time you get a refill. If you have any questions, ask your doctor or pharmacist.
This medication is best taken by mouth on an empty stomach, at least 1 hour before or 2 hours after a meal, usually 1 or 2 times daily or as directed by your doctor. Take this medication with a full glass of water (8 ounces/240 milliliters) unless directed otherwise. If stomach upset occurs, taking it with food or milk may help. However, doxycycline may not work as well if you take it with food or milk (or anything high in calcium - more details below ), so ask your doctor or pharmacist if you may take it that way. Do not lie down for at least 10 minutes after taking this medication.
Take this medication 2 to 3 hours before or after taking any products containing aluminum, calcium, iron, magnesium, zinc, or bismuth subsalicylate. Some examples include antacids, didanosine solution, quinapril, vitamins/minerals, dairy products (such as milk, yogurt), and calcium-enriched juice. These products bind with doxycycline, preventing your body from fully absorbing the drug.
When using to prevent malaria, this medication is usually taken once daily. Take the first dose of this medication 1 to 2 days before travel or as directed by your doctor. Continue to take this medication daily while in the malarious area. Upon returning home, you should keep taking this medication for 4 more weeks. If you are unable to finish this course of doxycycline, contact your doctor.
If you are using the liquid form of this medication, shake the bottle well before each dose. Carefully measure the dose using a special measuring device/spoon. Do not use a household spoon because you may not get the correct dose.
The dosage is based on your medical condition and response to treatment. For children, the dosage may also be based on weight.
For the best effect, take this antibiotic at evenly spaced times. To help you remember, take this medication at the same time(s) every day.
Continue to take this medication until the full prescribed amount is finished, even if symptoms disappear after a few days. Stopping the medication too early may allow bacteria to continue to grow, which may result in a return of the infection.
Tell your doctor if your condition persists or worsens.
Side Effects
Stomach upset, diarrhea, nausea, or vomiting may occur. If any of these effects persist or worsen, tell your doctor or pharmacist promptly.
Remember that this medication has been prescribed because your doctor has judged that the benefit to you is greater than the risk of side effects. Many people using this medication do not have serious side effects.
Tell your doctor right away if you have any serious side effects, including: painful/difficult swallowing, signs of kidney problems (such as change in the amount of urine).
Tetracycline drugs such as doxycycline may rarely cause a serious increase in pressure inside the skull (intracranial hypertension-IH). The risk of this side effect is greater for women of childbearing age who are overweight or who have had IH in the past. If IH develops, it usually goes away after doxycycline is stopped; however, there is a chance of permanent vision loss or blindness. Get medical help right away if you have: persistent/severe headache, vision changes (such as blurred/double vision, decreased vision, sudden blindness), persistent nausea/vomiting.
This medication may rarely cause a severe intestinal condition due to a bacteria called C. difficile. This condition may occur during treatment or weeks to months after treatment has stopped. Tell your doctor right away if you develop: diarrhea that doesn't stop, abdominal or stomach pain/cramping, blood/mucus in your stool.
If you have these symptoms, do not use anti-diarrhea or opioid products because they may make symptoms worse.
Use of this medication for prolonged or repeated periods may result in oral thrush or a new vaginal yeast infection. Contact your doctor if you notice white patches in your mouth, a change in vaginal discharge, or other new symptoms.
A very serious allergic reaction to this drug is rare. However, get medical help right away if you notice any symptoms of a serious allergic reaction, including: fever that doesn't go away, new or worsening lymph node swelling, rash, itching/swelling (especially of the face/tongue/throat), severe dizziness, trouble breathing.
This is not a complete list of possible side effects. If you notice other effects not listed above, contact your doctor or pharmacist.
Precautions
See also Side Effects section.
Before taking doxycycline, tell your doctor or pharmacist if you are allergic to it; or to other tetracyclines (such as minocycline); or if you have any other allergies. This product may contain inactive ingredients (such as sulfites, soy found in some brands), which can cause allergic reactions or other problems. Talk to your pharmacist for more details.
Before using this medication, tell your doctor or pharmacist your medical history, especially of: trouble swallowing, esophagus problems (such as hiatal hernia or reflux/heartburn).
Doxycycline may cause live bacterial vaccines (such as typhoid vaccine) to not work well. Tell your health care professional that you are using doxycycline before having any immunizations/vaccinations.
Before having surgery, tell your doctor or dentist that you are taking this medication.
This medication may make you more sensitive to the sun. Limit your time in the sun. Avoid tanning booths and sunlamps. Use sunscreen and wear protective clothing when outdoors. Tell your doctor right away if you get sunburned or have skin blisters/redness.
Children younger than 8 years may be more sensitive to the side effects of doxycycline, especially tooth discoloration. Tooth discoloration has also occurred in older children and young adults. Discuss the risks and benefits of this medication with the doctor.
Tell your doctor if you are pregnant or plan to become pregnant. You should not become pregnant while using doxycycline. Doxycycline may harm an unborn baby. If you become pregnant, talk to your doctor right away about the risks and benefits of this medication.
This medication passes into breast milk. Consult your doctor before breast-feeding.
Interactions
Drug interactions may change how your medications work or increase your risk for serious side effects. This document does not contain all possible drug interactions. Keep a list of all the products you use (including prescription/nonprescription drugs and herbal products) and share it with your doctor and pharmacist. Do not start, stop, or change the dosage of any medicines without your doctor's approval.
Some products that may interact with this drug include: retinoid medications taken by mouth (such as acitretin, isotretinoin), barbiturates (such as phenobarbital), "blood thinners" (such as warfarin), digoxin, anti-seizure medications (such as phenytoin), strontium.
This medication may interfere with certain laboratory tests (including urine catecholamine levels), possibly causing false test results. Make sure laboratory personnel and all your doctors know you use this drug.
Overdose
If someone has overdosed and has serious symptoms such as passing out or trouble breathing, call 911. Otherwise, call a poison control center right away. US residents can call their local poison control center at 1-800-222-1222. Canada residents can call a provincial poison control center.
Notes
Do not share this medication with others.
If this medication is being used to prevent malaria, it is important to understand that it is still possible to get the disease even if you have used this medication. Avoid being bitten by mosquitoes when trying to prevent malaria. Avoid contact with mosquitoes, especially from dusk to dawn, by staying in well-screened areas, wearing protective clothing, and using insect repellent and bed nets.
This medication has been prescribed for your current condition only. Do not use it later for another infection unless your doctor tells you to.
Laboratory and/or medical tests (such as complete blood count, liver/kidney function) may be performed periodically to monitor your progress or check for side effects. Consult your doctor for more details.
Missed Dose
If you miss a dose, take it as soon as you remember. If it is near the time of the next dose, skip the missed dose. Take your next dose at the regular time. Do not double the dose to catch up.
Storage
Store at room temperature away from light and moisture. Different brands of this medication have different storage needs. Check the product package for instructions on how to store your brand, or ask your pharmacist. Do not store in the bathroom. Keep all medications away from children and pets.
Do not flush medications down the toilet or pour them into a drain unless instructed to do so. Properly discard this product when it is expired or no longer needed. Taking outdated tetracycline-related drugs can result in serious illness. Consult your pharmacist or local waste disposal company.
Conclusion
As these studies have demonstrated both the antiviral and anti-cancer efficacy of doxycycline, with an excellent long established safety profile (subject to the above precautions), low cost and high availability it should be considered for inclusion in treatment regimes for Covid-19 and other viral infections and also for cancer patients.
Further to this, as it is not only compatible but indicated to work synergistically with both lactoferrin and/or ivermectin it should strongly be considered for inclusion in multi-drug clinical trials, albeit in a stepwise fashion and at a minimal dose to start with.
The only caution being that it should not be taken prophylactically due to the risk of inducing further bacterial and viral resistance.
DoorlessCarp’s Scientific Literature Reviews is a reader-supported publication. To receive new posts and support my work, consider becoming a free or paid subscriber.
Appendix
Hat tip to Nehming Names @ShemNehm on Gab.
From 2021, Rana et al conducted A Computational Study of Ivermectin and Doxycycline Combination Drug Against SARS-CoV-2 Infection (preprint)18
As well as ivermectin’s positive synergy with doxycycline the reverse has also been detected in silico, due to different molecular interactions:
“Molecular Dynamics study suggesting a synergism between Ivermectin and doxycycline, namely that together they bind more strongly to Covid's 3CL protease, and thus better incapacitate the virus.”
Abstract
In the present study, we have described how by using molecular docking and molecular dynamics (MD) simulation studies the combination drug of ivermectin and doxycycline can be used as a potential inhibitor for Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) virus. In lieu of unavailability of specific cure of coronavirus disease of 2019 (COVID-19) till now various possibilities for individual and combination drugs have been explored by the medical practitioners/scientists for the remedial purpose of CoV-2 infections. 3C-like protease (3CLpro) is the main protease of SARS-CoV-2 virus which plays an essential role in mediating viral replication in the human body. 3CLpro protein can serve as an attractive drug target. In this work, we have studied drug: 3CLpro interactions by in-silico molecular docking and MD simulation approaches. Common and easily available antiviral drugs ivermectin, doxycycline and their combination can regulate 3CLpro protein's function due to its easy inhibition.
A big thank you to Hedley Rees for contributing this excellent reference for health professionals: What’s stopping doctors repurposing generic medicines? Answer - not a lot!
Not to say this would be an easy accomplishment by any means, but the great thing about doxycycline is that it would come under the classification of “small-molecule” and it has a long and detailed history of regulatory approval.
Added 23rd December ‘23:
Caution is warranted as doxycycline is contraindicated for some patients, which is by no means unusual:
Doxycycline Promotes Carcinogenesis & Metastasis via Chronic Inflammatory Pathway: An In Vivo Approach (2016)
This site is strictly an information website about potential therapeutic agents and a review of the current state of research. It does not advertise anything, provide medical advice, diagnosis or treatment. This site is not promoting any of these as potential treatments or offers any claims for efficacy. Its content is aimed at researchers, registered medical practitioners, nurses or pharmacists. This content is not intended to be a substitute for professional medical advice, diagnosis, or treatment. Always seek the advice of your physician or other qualified health provider with any questions you may have regarding a medical condition. Never disregard professional medical advice or delay in seeking it because of something you have read on this website. Always consult a qualified health provider before introducing or stopping any medications as any possible drug interactions or effects will need to be considered.
ME Faure, A Cilibrizzi, V Abbate, KD Bruce, RC Hider. Effect of iron chelation on anti-pseudomonal activity of doxycycline, International Journal of Antimicrobial Agents, Volume 58, Issue 6, (2021), 106438, ISSN 0924-8579,
Alkawash M, Head M, Alshami I, Soothill JS. The effect of human lactoferrin on the MICs of doxycycline and rifampicin for Burkholderia cepacia and Pseudomonas aeruginosa strains. J Antimicrob Chemother. 1999 Sep;44(3):385-7. doi: 10.1093/jac/44.3.385. PMID: 10511407.
Wu ZC, Wang X, Wei JC, Li BB, Shao DH, Li YM, Liu K, Shi YY, Zhou B, Qiu YF, Ma ZY. Antiviral activity of doxycycline against vesicular stomatitis virus in vitro. FEMS Microbiol Lett. 2015 Nov;362(22):fnv195. doi: 10.1093/femsle/fnv195. Epub 2015 Oct 12. PMID: 26459887.
Mosquera-Sulbaran JA, Hernández-Fonseca H. Tetracycline and viruses: a possible treatment for COVID-19? Arch Virol. 2021 Jan;166(1):1-7. doi: 10.1007/s00705-020-04860-8. Epub 2020 Nov 2. PMID: 33136210; PMCID: PMC7604546.
Mahmud R, Rahman MM, Alam I, Ahmed KGU, Kabir AKMH, Sayeed SKJB, Rassel MA, Monayem FB, Islam MS, Islam MM, Barshan AD, Hoque MM, Mallik MU, Yusuf MA, Hossain MZ. Ivermectin in combination with doxycycline for treating COVID-19 symptoms: a randomized trial. J Int Med Res. 2021 May;49(5):3000605211013550. doi: 10.1177/03000605211013550. PMID: 33983065; PMCID: PMC8127799.
Buss JL, Torti FM, Torti SV. The role of iron chelation in cancer therapy. Curr Med Chem. 2003 Jun;10(12):1021-34. doi: 10.2174/0929867033457638. PMID: 12678674.
Richardson A, Kovacevic Z, Richardson DR. Iron chelation: inhibition of key signaling pathways in the induction of the epithelial mesenchymal transition in pancreatic cancer and other tumors. Crit Rev Oncog. 2013;18(5):409-34. doi: 10.1615/critrevoncog.2013007921. PMID: 23879587.
Morales M, Xue X. Targeting iron metabolism in cancer therapy. Theranostics. 2021 Jul 25;11(17):8412-8429. doi: 10.7150/thno.59092. PMID: 34373750; PMCID: PMC8344014.
Ibuki M, Shoda C, Miwa Y, Ishida A, Tsubota K, Kurihara T. Lactoferrin Has a Therapeutic Effect via HIF Inhibition in a Murine Model of Choroidal Neovascularization. Front Pharmacol. 2020 Feb 28;11:174. doi: 10.3389/fphar.2020.00174. PMID: 32180725; PMCID: PMC7059857.
Fife RS, Sledge GW Jr, Roth BJ, Proctor C. Effects of doxycycline on human prostate cancer cells in vitro. Cancer Lett. 1998 May 15;127(1-2):37-41. doi: 10.1016/s0304-3835(98)00003-2. PMID: 9619856.
Zhu C, Yan X, Yu A, Wang Y. Doxycycline synergizes with doxorubicin to inhibit the proliferation of castration-resistant prostate cancer cells. Acta Biochim Biophys Sin (Shanghai). 2017 Nov 1;49(11):999-1007. doi: 10.1093/abbs/gmx097. PMID: 28985240.
Markowska A, Kaysiewicz J, Markowska J, Huczyński A. Doxycycline, salinomycin, monensin and ivermectin repositioned as cancer drugs. Bioorg Med Chem Lett. 2019 Jul 1;29(13):1549-1554. doi: 10.1016/j.bmcl.2019.04.045. Epub 2019 Apr 27. PMID: 31054863.
Dr. Meenakshi Rana, Pooja Yadav, Papia Chowdhury et al. A Computational Study of Ivermectin and Doxycycline Combination Drug Against SARS-CoV-2 Infection, 05 August 2021, PREPRINT (Version 1) available at Research Square [https://doi.org/10.21203/rs.3.rs-755838/v1]
For bone mets, breast cancer patients have long been advised to take doxy.
My wariness with doxy as being a member of the tetracycline family is: "Tetracyclines, Even at low concentrations, tetracyclines induce mitochondrial proteotoxic stress, leading to changes in nuclear gene expression and altered mitochondrial dynamics and function in commonly used cell types, as well as worms, flies, mice, and plants. "
Mitochondria, of course, are comprised of vestigial bacteria. Are we destroying our mitochondria by doxy's attack on mitochondria? I am passionate about helping cancer patients, especially breast cancer patients, and I have always passed along this caveat.










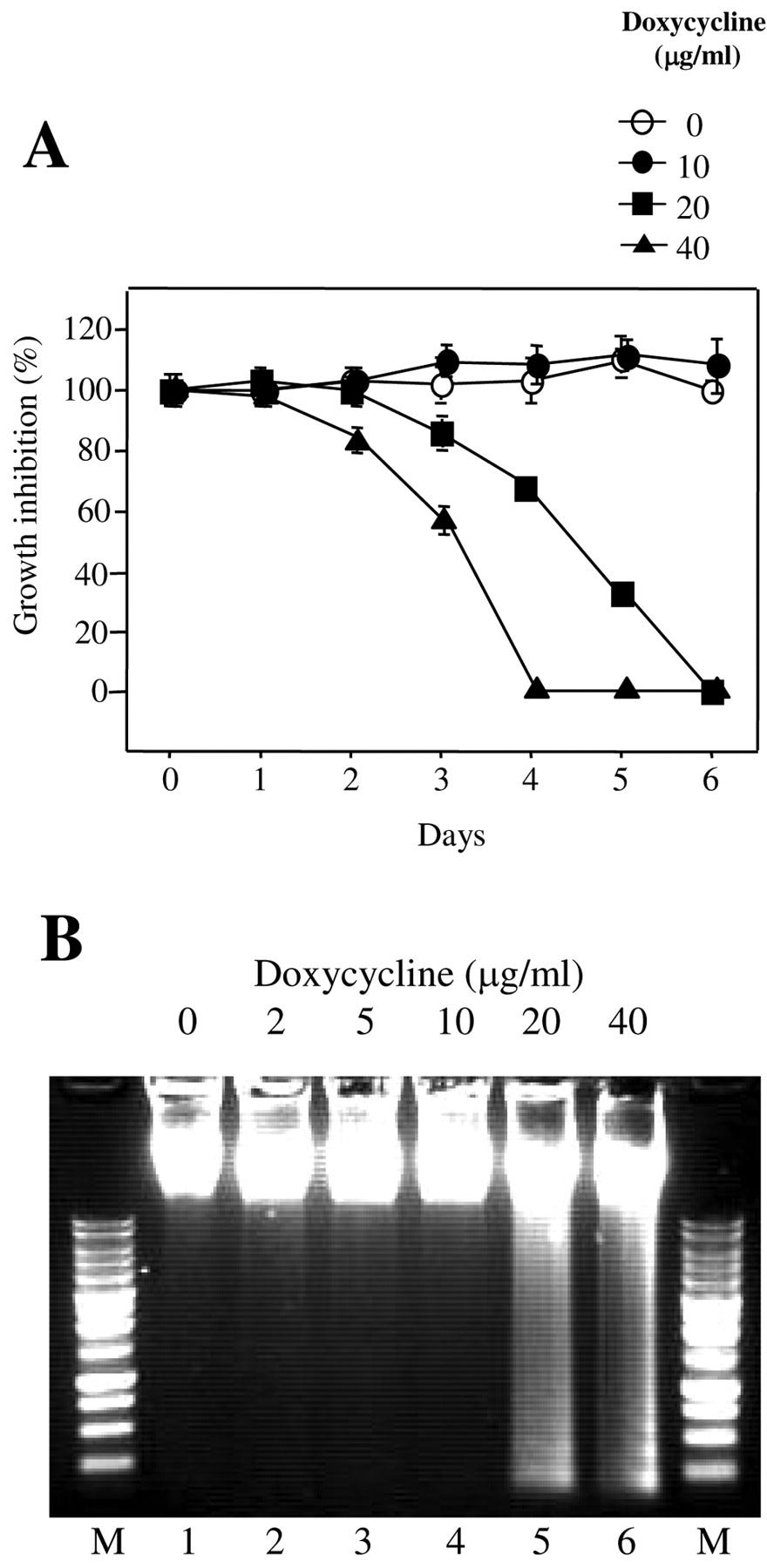
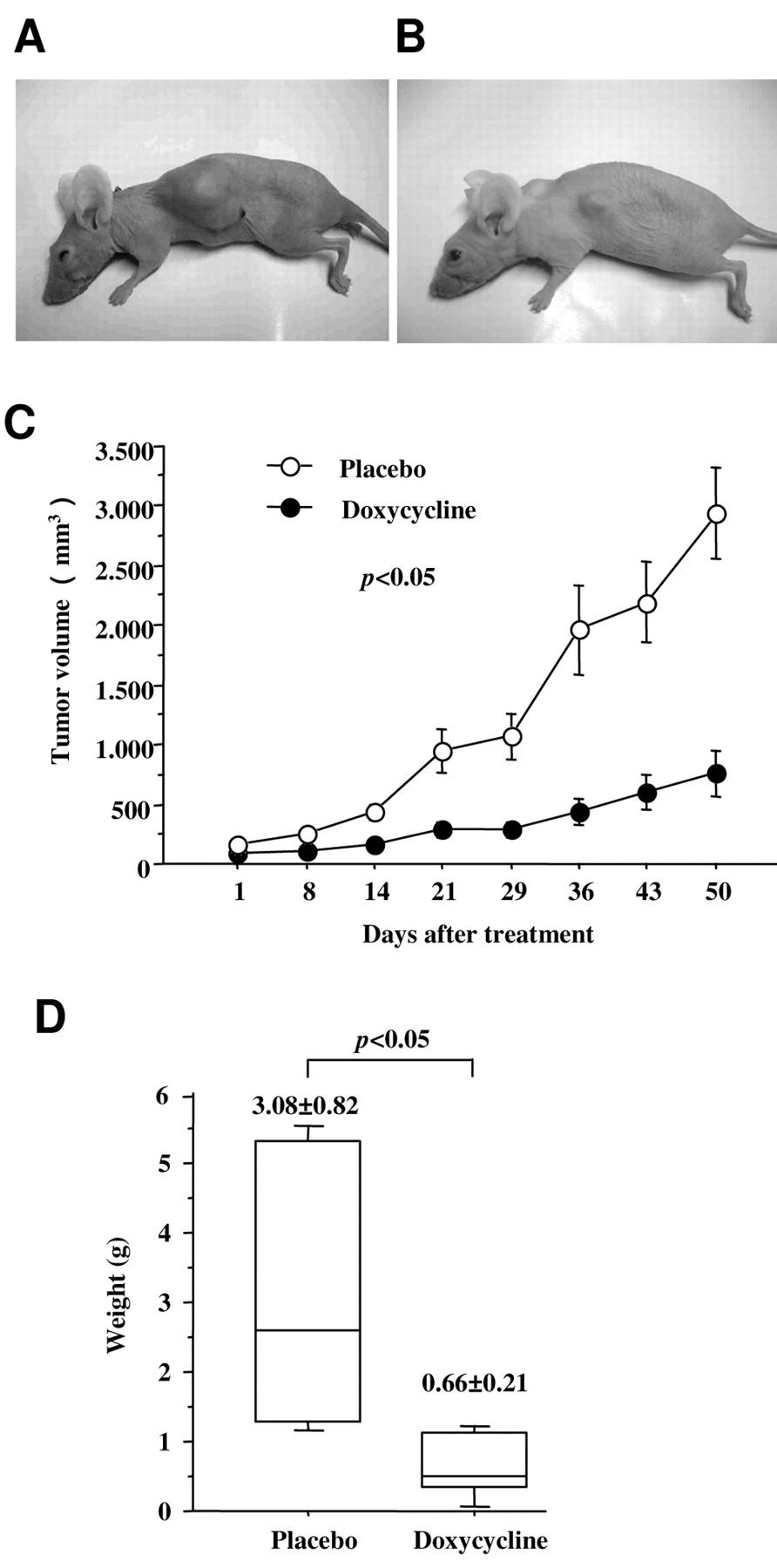






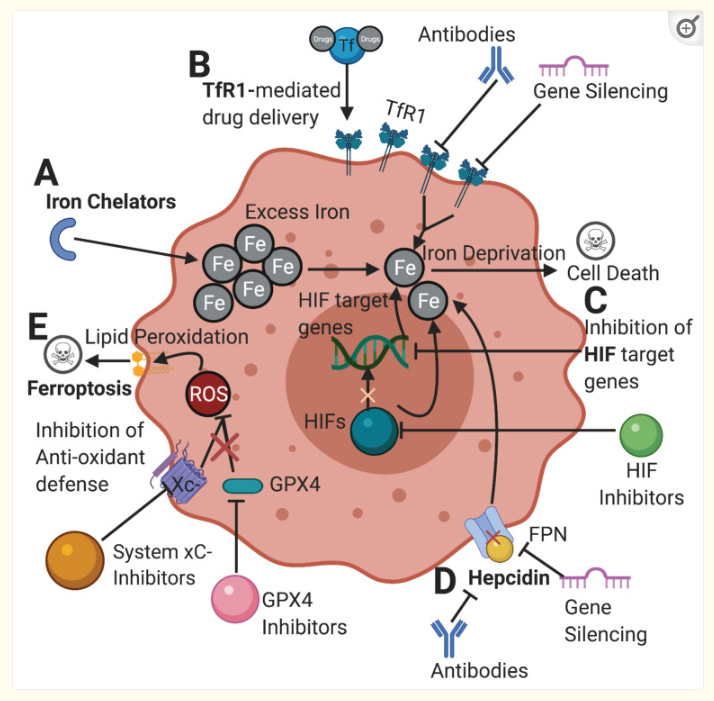
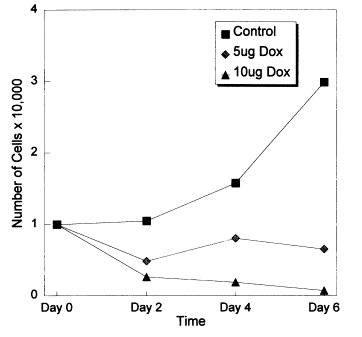





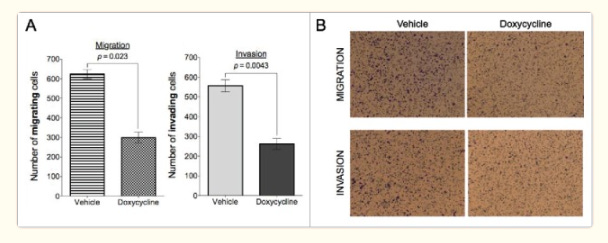
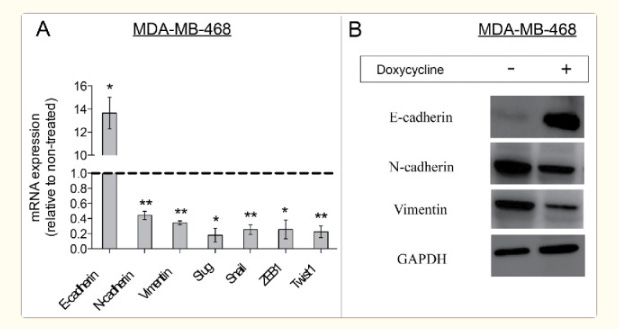






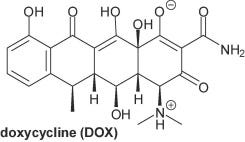




As always, thank you, Doorless Carp!
For bone mets, breast cancer patients have long been advised to take doxy.
My wariness with doxy as being a member of the tetracycline family is: "Tetracyclines, Even at low concentrations, tetracyclines induce mitochondrial proteotoxic stress, leading to changes in nuclear gene expression and altered mitochondrial dynamics and function in commonly used cell types, as well as worms, flies, mice, and plants. "
Mitochondria, of course, are comprised of vestigial bacteria. Are we destroying our mitochondria by doxy's attack on mitochondria? I am passionate about helping cancer patients, especially breast cancer patients, and I have always passed along this caveat.
THANKS AGAIN, DOORLESS CARP!
Thanks for this Mr Carp! Readers may find this if interested, written when I was helping Dr Tess Lawrie on IVM approval by MHRA: https://www.dropbox.com/s/4ygwnz4o76exd5i/GMP%20Vol20No2_Rees.pdf?dl=0