Any extracts used in the following article are for non commercial research and educational purposes only and may be subject to copyright from their respective owners.
The kynurenine pathway is a metabolic pathway leading to the production of nicotinamide adenine dinucleotide (NAD+). Metabolites involved in the kynurenine pathway include tryptophan, kynurenine, kynurenic acid, xanthurenic acid, quinolinic acid, and 3-hydroxykynurenine. The kynurenine pathway is responsible for total catabolization of tryptophan about 95%. Disruption in the pathway is associated with certain genetic and psychiatric disorders.
Kynurenine pathway dysfunction
Disorders affecting the kynurenine pathway may be primary (of genetic origin) or secondary (due to inflammatory conditions). Peripheral inflammation can lead to a build up of kynurenine in the brain, and this is associated with major depressive disorder, bipolar disorder, and schizophrenia. Dysfunction of the pathway not only cause increasing metabolites such as quinolinic acid or kynurenic acid but also affects serotonin and melatonin synthesis. Kynurenine clearance in exercised muscle cells can suppress the build up in the brain.
Dysfunctional states of distinct steps of the kynurenine pathway (such as kynurenine, kynurenic acid, quinolinic acid, anthranilic acid, 3-hydroxykynurenine) have been described for a number of disorders, for example:
HIV dementia
Tourette syndrome
Tic disorders
Psychiatric disorders (such as schizophrenia, major depressive disorder, bipolar disorder, anxiety disorders)
Multiple sclerosis
Huntington's disease
Encephalopathies
Lipid metabolism
Liver fat metabolism
Systemic lupus erythematosus
Glutaric aciduria
Vitamin B6 deficiency
Eosinophilia-myalgia syndrome
long COVID
Tryptophan (symbol Trp or W) is an α-amino acid that is used in the biosynthesis of proteins. Tryptophan contains an α-amino group, an α-carboxylic acid group, and a side chain indole, making it a polar molecule with a non-polar aromatic beta carbon substituent. It is essential in humans, meaning that the body cannot synthesize it and it must be obtained from the diet. Tryptophan is also a precursor to the neurotransmitter serotonin, the hormone melatonin, and vitamin B3. It is encoded by the codon UGG.
Like other amino acids, tryptophan is a zwitterion at physiological pH where the amino group is protonated (–NH+3; pKa = 9.39) and the carboxylic acid is deprotonated ( –COO−; pKa = 2.38).[5]
Humans and many animals cannot synthesize tryptophan: they need to obtain it through their diet, making it an essential amino acid
Dietary sources
Tryptophan is present in most protein-based foods or dietary proteins. It is particularly plentiful in chocolate, oats, dried dates, milk, yogurt, cottage cheese, red meat, eggs, fish, poultry, sesame, chickpeas, almonds, sunflower seeds, pumpkin seeds, Hemp Seeds, buckwheat, spirulina, and peanuts. Contrary to the popular belief that cooked turkey contains an abundance of tryptophan, the tryptophan content in turkey is typical of poultry.1
Pathophysiology
Indoleamine-pyrrole 2,3-dioxygenase (IDO or INDO EC 1.13.11.52) is a heme-containing enzyme physiologically expressed in a number of tissues and cells, such as the small intestine, lungs, female genital tract or placenta. In humans is encoded by the IDO1gene. IDO is involved in tryptophan metabolism. It is one of three enzymes that catalyze the first and rate-limiting step in the kynurenine pathway, the O2-dependent oxidation of L-tryptophan to N-formylkynurenine, the others being indolamine-2,3-dioxygenase 2 (IDO2) and tryptophan 2,3-dioxygenase (TDO). IDO is an important part of the immune system and plays a part in natural defense against various pathogens. It is produced by the cells in response to inflammation and has an immunosuppressive function because of its ability to limit T-cell function and engage mechanisms of immune tolerance. Emerging evidence suggests that IDO becomes activated during tumor development, helping malignant cells escape eradication by the immune system. Expression of IDO has been described in a number of types of cancer, such as acute myeloid leukemia, ovarian cancer or colorectal cancer. IDO is part of the malignant transformation process and plays a key role in suppressing the anti-tumor immune response in the body, so inhibiting it could increase the effect of chemotherapy as well as other immunotherapeutic protocols.2
In this paper from 2016, Routy et al discuss how the kynurenine pathway (KP) is a double edged sword: both protecting some tissues and organs from autoimmune pathology & the effects of inflammatory cytokines on the one hand, but also permitting tumor growth and metastasis on the other.
Abstract
The term "immune privilege" was originally coined to describe the suppression of inflammatory responses within organs protected by anatomic barriers, ie, the eyes, brain, placenta, and testes. However, cellular and metabolic processes, which orchestrate immune responses, also control inflammation within these sites. Our current understanding of tolerogenic mechanisms has extended the definition of immune privilege to include hair follicles, the colon, and cancer. By catabolizing tryptophan, cells expressing the enzyme indoleamine-2,3-dioxygenase produce kynurenine metabolites, which orchestrate local and systemic responses to control inflammation, thus maintaining immune privilege. This review highlights the double-edged role played by the kynurenine pathway (KP), which establishes and maintains immune-privileged sites while contributing to cancer immune escape. The identification of the underlying molecular drivers of the KP in immune-privileged sites and in cancer is essential for the development of novel therapies to treat autoimmunity and cancer and to improve transplantation outcomes.
The KP contributes to immune privilege by the following four mechanisms: (1) by depleting Trp via the induction of the stress response kinase, general control nondepressible 2 (GCN2), and the suppression of mammalian target of rapamycin 1 (mTOR1) pathway, which senses amino acid withdrawal, a process inhibiting Teff cell function and growth (Fig. 2)33,34; (2) by the direct effect of Kyn on the aryl hydrocarbon receptor (AhR), which induces DC and MP differentiation; these cells in turn first induce inflammatory Th17 cells followed by a global reprogramming driving their conversion (transdifferentiation) to Treg cells upon the resolution of inflammation.35 In the context of chronic inflammation, the effect of Kyn on AhR is directed toward a predominant immune suppression; (3) by promoting the differentiation of CD4 T cells into Treg cells expressing CTLA-4 and via phosphatase and tensin homolog, a protein encoded by a tumor suppressor gene, which is mutated in several cancers36; and finally (4) by the Kyn-mediated inhibition of IL-2 signaling, which impairs memory CD4 T cell survival.37–39Upon IDO activation, APCs capable of producing inflammatory cytokines such as IL-12 switch to produce inhibitory cytokines such as transforming growth factor beta (TGF-β) and IL−10.37In addition, IDO can be triggered in the presence of apoptotic cells to induce self-tolerance and/or cancer persistence.40 Globally, IDO activation can transform the function of APCs and convert local T cell function from an immunogenic one to a tolerogenic one.
Figure 2 The role of the kynurenine pathway in inducing tolerance in immune-privileged sites, in tumors, and in the tumor microenvironment.
Notes: The production of kynurenine by the IDO enzymatic activity of tumor cells and of APCs leads to immune tolerance in immune-privileged sites and in the tumor microenvironment. Kyn plays a major role by inducing Treg cells, which can also be directly induced by the cytokines and chemokines produced by tumor cells. The cytokines and chemokines produced by tumor cells can also stimulate APCs to activate the KP, which contributes further to tumor development. The four major mechanisms of the KP that influence the immune response are as follows: (1) Trp depletion followed by GCN2 induction and mTOR1 suppression; (2) activation of AhR by Kyn; (3) Treg induction through expression of CTLA-4 and PTEN; and (4) Kyn-mediated blockade of IL-2. Other factors implicated in immune tolerance include tumor cell expression of PD-1/PD-L1 and macrophage expression of MIF. Dotted lines represent decreased Trp concentrations.
Cancer and its Microenvironment: Kyn is the Enemy Within
In 2008, Mellor and Munn10 extended the definition of immune privilege to include cancer and its microenvironment. In the early stage of tumor development, proteins encoded by mutated genes are thought to be the major source of antigens in cancer cells, which are able to circumvent tumor-specific immune responses owing to the immune-privileged tumor or its microenvironment.15,100,101
IDO can be expressed by tumor cells and by myeloid cells surrounding the tumor; these myeloid cells include APCs, tumor-associated macrophages (TAMs), and myeloid-derived suppressor cells (MDSCs), which mediate an acquired immune tolerance toward the tumor, thereby thwarting host immune responses.It was recently recognized that the tumor’s metabolic microenvironment plays a major role in maintaining immune responses by causing a shift from mitochondrial oxidative phosphorylation, an efficient way to generate ATP, to a less efficient aerobic glycolysis pathway in cytotoxic T cells (CTLs).102 This metabolic reprogramming or Warburg effect within the tumor and its microenvironment leads initially to a local immune exhaustion involving CTLs and NK cells, which will become systemic when metastasis develops. Such cancer-related immune exhaustion has recently been shown to be influenced by the gut microbiota in a distal nondigestive tumor.103 Furthermore, cytokines and chemokines secreted by the hypoxic and necrotic zones of the tumor microenvironment attract TAMs; the presence of high numbers of TAMs expressing high levels of IDO and arginase-1 is a phenomenon linked to immune escape and to faster disease progression.
The tumor microenvironment also accumulates immature and tolerogenic tumor-infiltrating APCs expressing low levels of costimulatory proteins such as CD80 and CD86; thus, these APCs lack the ability to stimulate naive T cells.102The migration of these APCs into the draining lymph nodes in which the APCs present tumor antigens to tumor-specific T cells subsequently results in theirfailure to activate T cells. In mice, MDSCs infiltrate the tumor microenvironment and promote T cell dysfunction and Treg cell generation through the expression of IDO and arginase-1.104 Targeting MDSCs with monoclonal antibodies has been shown to restore the tumoricidal function of tumor-infiltrating T lymphocytes in mice.
In 2003, Uyttenhove et al.105 were the first to describe the constitutive expression of IDO by either the tumor cells or the APCs in the tumor microenvironment of various human cancers including cancers of the lung, pancreas, stomach, prostate, endometrium, and ovaries. They also showed that the expression of IDO by immunogenic murine tumor cells prevented their rejection by preimmunized mice, thereby inducing immune privilege. This effect was associated with the inhibition of specific T cell responses at the tumor site, an effect that was partly reversed by the administration of an IDO inhibitor. Subsequently, a number of studies reported an important role for the KP in several other cancers.106–109
Another immunometabolic pathway involving adenosine and ATP was recently described.85,110 Pericellular adenosine in the tumor microenvironment suppresses antitumor T cells by binding to the adenosine A2 A receptor, which activates cyclic adenosine monophosphate/protein kinase A signaling; this pathway subsequently inhibits IL-2 production in T cells, thereby suppressing the function of Teff cells. Interestingly, we have recently shown in vitro that a physiological concentration of Kyn (5 µM) was sufficient to reduce the ability of memory CD4 T cells to respond to IL-2,39 therefore identifying a mechanism that can be therapeutically targeted.
Other factors contributing to immune privilege within tumors include the following immune checkpoints and immunosuppressive cytokines: (1) immune checkpoints like PD-L1 and/or PD-L2, CTLA-4, and Fas, which are expressed at higher levels in the tumor and its microenvironment are the targets of several immune interventions; and (2) the influence of cytokines which was first reported in 1989 by Werner et al.111 who showed that the influence of IFN-γ on IDO expression in cell culture using various human cells. Litzenburger et al.112 established a link between AhR and the IL-6/STAT3 axis in driving IDO expression. However, Li et al.113 have shown that the long-term maintenance of IDO expression was found to be independent of IFN-γ, IL-10, TGF-β, TNF-α, and IL-6, contrasting with IDO enzymatic activity and IFN-γ-induced AhR expression. These immune checkpoint molecules and cytokines have been recently reviewed by Galluzzi and Kroemer,114 and their inhibition by therapeutic antibodies is under intense clinical evaluation.3
The importance of the gut biome (as above), COVID-19 severity and cancer risk as discussed by Cai et al (2021). This is in part due to the stimulation of inflammatory cytokines and the relationship between inflammation, the kynurenine pathway and carcinogenesis:
Previous studies showed that the condition of COVID-19 patients during hospitalization was closely related to the abundance of gut microbiota, and firmicutes might play an important role. Gou et al. found [10] that theBlautia genus, Lactobacillus genus, and Ruminococcus genus of Firmicutes in the gut microbiota of COVID-19 patients is correlated with the expression of multiple inflammatory molecules. Meanwhile, the operational taxonomic unit (OTU) composed of these three genera is correlated with the severity of COVID-19.
COVID-19 patients with cancer, including colorectal cancer (CRC), have a higher risk of severe events than their counterparts who do not have cancer, which may be associated with advancing age, male sex, but its relationship to gut microbiota need to be further researched [11, 12]. It is more difficult to treat COVID-19 patients with cancer than others because of their complex immune environment [13, 14], and gut microbiota may provide a new therapeutic avenue for these patients. Previous studies showed that the occurrence, development, and treatment of a variety of cancers, including colorectal cancer, are strongly related to gut microbiota [15,16,17,18]. The gut microbiota spectrum of colorectal cancer patients is significantly different from that of the healthy control group, with the increase of Fusobacterium nucleatum, Parvimonas, Peptostreptococcus, Porphyromonas, and Prevotella genera [19,20,21,22,23,24,25,26]. Also, several studies indicated that the abundance of Blautia [27, 28] and Ruminococcus [29, 30] changed in the gut microbiota of colorectal cancer patients, and changes in the abundance of these Microbes may affect the severity of COVID-19 disease. However, the correlation between the gut microbiota of tumor patients and COVID-19 remains to be studied.4
An interesting preprint, (version 3, May ‘22) by Luiz Zanella discusses the theory that some of the pathophysiology of COVID-19 is because SARS-CoV-2 mimics the Warburg Effect (WE) of cancer and that long COVID can be a WE. Tryptophan catabolism is part of the WE.
Abstract
This article develops the theory involving the SARS-CoV-2 pathophysiological mechanisms, based on an extensive literature review, and reinforced by two years of dealing with COVID-19 patients in different footages of the natural history of the disease. SARS-CoV-2 mimics the Warburg effect (W.E.), a well-described mechanism in neoplastic cells to obtain energy and substrates to enable cell growth. The W.E. is responsible for characteristic clinical and laboratory findings such as elevated lactic dehydrogenase (LDH), lactate, uraemia, and acidosis, in addition to explaining why a specific profile of patients evolves into severe forms of the disease. This article also exposes a hypothesis for maintaining the inflammatory status after acute SARS-COV-2 infection, "The Long COVID-19 Syndrome". Assessment of PubMed platform followed by evaluation of articles that supported issues regarding COVID-19/SARS-CoV-2 AND AEROBIC GLYCOLISIS/WARBURG EFFECT. COVID-19 is a biphasic infectious disease (like Yellow Fever, for example) that causes tenacious immune-metabolic changes that impact the inflammatory status. The hyperlactataemia (type B) in COVID-19 is due to the metabolic shift to aerobic glycolysis (W.E.), and it can differentiate chronic or acute COVID-19 inflammatory status from septic shock hyperlactatemia, especially when associated with negative cultures. This article brings to the light the pathophysiology of SARS-CoV-2 and possibilities for treatment and improvement in the clinical management of patients affected by this severe and catastrophic disease. The COVID-19 signature is the Warburg Effect + ACE-2 + innate response + hypoxia + Phenylalanine (Phe), and Tryptophan (Try) metabolism. The disease’s magnitude is directly proportional to the patient's exposure to hypoxia.
SARS-COV-2 uses the Warburg Effect to obtain energy and substrate for its replication. Other viruses also use this exact mechanism, usually via viral proteins that shift metabolism to aerobic glycolysis, to the detriment of oxidative phosphorylation (OXPHOS), which remains active, but is of lesser importance for the maintenance of viral replication mechanisms. EW is also used physiologically by some human body cells, such as the myocardium and cells of innate and adaptive immunity.
The variation between aerobic mechanisms of cellular respiration and aerobic glycolysis is orchestrated impacts the differentiation of immune system cells and the possibility of neoplastic cells to develop and cause metastasis(3–6).
The profile of patients who evolve to the most severe forms of COVID-19 (severe and critical) are those who have inflammatory comorbidities with some degree of hypoxia so that their cells already have their metabolic pathways diverted to perform W.E., and hyperlactatemia is due to metabolic deviation, although mixed situations, that is, increased lactate associated with shock conditions (septic shock, for example) can occur due to immunosuppression secondary to COVID-19(7,8).
COVID-19 presents a very variable chronic form, presenting itself in a pleomorphic form, making its diagnosis difficult. Although the chronic inflammatory status (called "The Long COVID-19 Syndrome) has multiple faces, some signs and symptoms such as gastrointestinal, airway, neuropsychiatric are prevalent. Inflammatory Shock (IS) is the most severe outcome in COVID-19, which can occur immediately after the acute phase of the disease or later (Weeks or months after acute disease). Inflammatory shock, often confused with septic shock and rarely diagnosed, causes great underreporting of deaths from COVID-19.
The Warburg Effect/SARS-CoV-2 relationship and the presented phenomena in the clinical and laboratory evaluation
The interference of SARS-CoV-2 in the human immunometabolic pathways seems to be the cause of the severity of the disease, especially in those who have inflammatory comorbidities and divert cellular metabolism to the glycolytic pathway and the immune system to create a tolerant environment in the presence of an inflammatory state. These changes are even more intensified and directly proportional to the lung injury caused by the coronavirus.
The immune axis influences the metabolic axis and vice versa.
By initiating the immune response mainly via the production of Interferon-gamma (IFN-𝛾), the axis of Try metabolism is diverted to the production of Kynurenine, negatively interfering with the serotonin and NAD/NADH + pathways, which are usually in the state of homeostasis. Try is processed by the enzyme Tryptophan hydroxylase (TDO) when in homeostasis; however, in the inflammation status, the liver stops producing TDO and the processing of Try starts to be mediated by the indoleamine 2,3-dioxygenase (IDO) enzyme produced mainly by dendritic cells (DCs), Macrophages (MO) and Monocytes. It remains controversial how IDO-1 influences the tendency towards the Kynurenine pathway. KYNA produces some toxic metabolites that bind to GABA receptors. These metabolites are found in diseases such as Alzheimer's and Parkinson's. These metabolites appear to exert significant neuropsychiatric symptoms in COVID-19 patients, whose symptoms are prevalent. In addition, they hinder the action of anaesthetics in sedation and intubation processes and activate pain mechanisms, especially peripheral ones(30–32).
The deviation of the Try axis promotes lower production of NAD/NADH due to low production of the B complex (mainly B3), causing Pellagra signs in patients and an enzymatic deficit in intracellular aerobic respiration. 5-HT is also compromised, producing less Melatonin, and causing acute depressive symptoms, even in patients with no previously reported psychiatric illness history(33–35).
Phe is mainly interfered with by neopterin produced mainly by Monocytes and DCs by IFN-𝛾 stimulation. In monocyte-derived macrophages and dendritic cells, IFN-𝛾 triggers GTP-cyclohydrolase-I, the key enzyme for pteridine derivatives biosynthesis like neopterin 5,6,7,8-tetrahydrobiopterin (BH4).
Neopterin has been linked to the mortality outcome in cardiovascular and other inflammatory diseases, in addition, it influences the increase in the Phe/Tyr and Kin/Tryp ratio PAH may be due to an increased output of reactive oxygen species (ROS), which is produced by macrophages upon stimulation by IFN-𝛾 in parallel to neopterin production(40–42).
The decrease in Tyr due to the increase in Phe/Tyr influences the decreased production of Melatonin, affecting the sleep-wake dynamics, with insomnia being a prevalent symptom in acute COVID-19 and the "Long COVID-19 Syndrome". Melatonin is synthesized from serotonin in a two-step process by the sequential action of the key enzymes aralkylamine N-acetyltransferase (AANAT) and acetylserotonin O-methyltransferase (ASMT)(36,43–46).
Melatonin acts in the human body as a sleep-inducing hormone and still preserves the evolutionary characteristic of interacting with various ROS to produce cyclic 3-hydroxymelatonin and other melatonin metabolites, e.g., N1-acetyl-N2-formyl-5-methoxykynuramine and N-acetyl-5-methoxykynuramine. These metabolites function as radical scavengers, sometimes even more aggressively than Melatonin, regarding their capacity to neutralize ROS. Melatonin stimulates the antioxidant enzyme superoxide dismutase (SOD2), an action that involves an elevated level of sirtuin 3 (SIRT3). Melatonin enters mitochondria through the oligopeptide transporters, PEPT1/2 and influences mitochondrial membrane potential by uncoupling protein(UCP); finally, Melatonin from the matrix may leak out of the mitochondria to interact with the melatonin receptors, MT1 and MT2, to control the release of cytochrome c(47–49).
The Warburg effect, also called aerobic glycolysis, is a mechanism in which the cell obtains energy (ATP) through the glycolytic pathway at the expense of oxidative phosphorylation and is initially described for neoplastic cells hypoxemic environments with and without hyperglycaemia(3,4,66–69).
The hyperglycaemic environment favours the development of the Warburg effect, as hypoxia and inflammation intensify this effect. That is why inflammatory and neoplastic diseases, obesity, 2MDD and advanced heart failure (diseases now considered inflammatory because they promote tissue hypoxia and, consequently, inflammation) are predictors of severity in COVID-19.
What differentiates neoplastic wasting syndrome from "The long COVID-19 syndrome" is the fact that the latter has acute weight loss, in addition to acute delirium with acute loss of functionality, elevated LDH and lactate and no previous history of neoplasia. Although COVID-19 may open the door to the rapid development of neoplastic cells due to cellular changes brought about by oxidative stress and W.E. (6,69,72,73)
W.E. also acts on the immune system, favouring a more tolerant environment than the inflammatory one, being responsible, for example, in the occurrence of metastases, considering its effect on neoplastic cells. The detailed mechanisms of EW are covered in the "APPENDIX" of this article(3,81–86).
W.E. acts on the immune system in several ways, tending the immune system towards tolerance. Negative signalling induced by ligand binding causes a downregulation of the immune response to avoid overactivation of immune activities, and some of the immune checkpoints have been identified, such as PD-1, CTLA4, TIM3, and so on. W.E. stimulates macrophage polarization to M2 induces apoptosis of NK and T-cells of the inflammatory spectrum while favouring innate immunity and the Treg pole.
COVID-19 is a multisystem infectious disease since the ACE-2 receptor, for a viral infection to occur, is disseminated to almost all human body tissues.
The most recent proteomics studies show alterations in the metabolism of lipids, Branched-chain amino acids (BCAAs), Tryptophan, Phenylalanine, and components of the citric acid cycle, probably related to the shift from oxidative metabolism, mediated by OXPHOS to aerobic glycolysis metabolism, called the Warburg effect(72,78).
Many viruses use W.E. to carry out replication more effectively within cells, as aerobic glycolysis provides substrate (nitrogenous bases and ATP) for producing genetic material and molecules that make up the viral organization. W.E. is used by neoplastic cells for growth and metastasis and usually occurs in a hyperglycaemic and hypoxemic environment. Although OXPHOS continues to occur, it is less used as the metabolic changes that occur trigger a truncated citric acid cycle.
COVID-19 patients, to a greater or lesser extent, undergo a process of immunosuppression resulting from W.E., although they remain, at the same time, with a tendency to activate the TH17 immune response. This inflammatory environment concomitant with the shift from the pole to Treg is conducive to the emergence of autoantibodies. In addition, lymphocytic apoptosis contributes to the development of neoplasms since there is the death of N.K. and TCD8 + in an environment where cells are under the significant influence of oxidative stress, altering all cellular control mechanisms. Many prevalence studies are still needed, but the development of neoplastic diseases (mainly breast and gastrointestinal system) linked to previous SARS-CoV-2 infection is already being noticed.5
Gabrieli et al (2017) on the relationship between the catabolic products of tryptophan and the metabolism & malignancy of a type of brain cancer with a very poor survival rate, glioblastoma (GBM):
Trends
Metabolic remodeling in tumor and immune cells promotes GBM malignancy.
In addition to the well-characterized Warburg effect, tryptophan catabolism contributes to gliomagenesis through a kynurenine–AHR signaling pathway.
HIF-1α and AHR sense metabolic changes in the tumor environment and promote glioma progression by signaling in not only cancer, but also immune cells, such as CD4 and CD8 T lymphocytes and macrophages.
Glioblastoma (GBM) progression is associated with metabolic remodeling in both glioma and immune cells, resulting in the use of aerobic glycolysis as the main source of energy and biosynthetic molecules. The transcription factor hypoxia-inducible factor (HIF)-1α drives this metabolic reorganization. Oxygen levels, as well as other factors, control the activity of HIF-1α. In addition, the ligand-activated transcription factor aryl hydrocarbon receptor (AHR) modulates tumor-specific immunity and can also participate in metabolic remodeling. AHR activity is regulated by tryptophan derivatives present in the tumor microenvironment. Thus, the tumor microenvironment and signaling via HIF-1α and AHR regulate the metabolism of gliomas and immune cells, modulating tumor-specific immunity and, consequently, tumor growth. Here, we review the roles of HIF-1α and AHR in cancer and immune cell metabolism in GBM.
Figure 1. Factors in the tumor microenvironment drive immunosuppression in GBM Soluble factors in the periphery and tumor microenvironment can shift adaptive and innate immune cell function to an immunosuppressive state. Microbiota products and onco-metabolites provide AHR agonists that inhibit inflammatory cytokine production by dendritic cells and macrophages, favoring the differentiation of CD4+ Treg cells. Moreover, hypoxic microenvironments favor tumor growth by inducing pro-tumorigenic macrophage polarization.
The malignancy of GBM is manifested by its high proliferation rate, ability to invade the surrounding brain parenchyma and immunosuppression. These aspects of GBM malignancy are supported by the manipulation of a number of biological pathways to exploit not only intracellular tumor resources but also the microenvironment provided by surrounding cells [28]. Augmented glycolysis or Warburg effect [3, 28, 29] and abnormal tryptophan catabolism [30] are hallmarks of GBM. Specifically, Xiong and colleagues observed that a mutated form of a critical component of the TCA cycle, isocitrate dehydrogenase 1 (IDH1), is associated with HIF-1α-mediated carcinogenesis. Their findings provide an important line of evidence that links metabolic dysfunction, through disruption of the IDH1 pathway and increases in HIF-1α activity, to increased transcriptional activity leading to more aggressive glioma growth.
In addition, Michael Platten’s group reported that kynurenine, an endogenous ligand for the transcription factor AHR derived from tryptophan is produced by glioma cells. The group suggested that the kynurenine-AHR pathway contributes to GBM pathology by increasing the growth and motility of tumor cells and suppressing the immune response. Therefore, to understand the role of metabolism in GBM pathology it is important to link specific metabolic signaling pathways to specific cell populations in the tumor microenvironment.
Multiple lines of evidence indicate that the increase in aerobic glycolysis detected in GBM supports the elevated nutrient demands of fast proliferating cancer cells by providing lipid and nucleotide biosynthesis [28]. In addition, this increased glycolysis has important effects on tumor-specific immunity and consequently, tumor pathogenesis [31, 32]. Indeed, the Warburg effect also promotes the production of lactate in anaerobic conditions, which attracts immune cells to the tumor microenvironment where they are imprinted with a tumor suppressing phenotype [33–35]. For example, excessive accumulation of lactic acid in the tumor microenvironment can lead to disruption of the lactic acid gradient between the intracellular space of lymphoid cells and the extracellular milieu [21]. Through this mechanism, T-cells can no longer export intracellular lactic acid efficiently, which leads to a disruption of metabolic processes and a consequent decrease in T-cell function. Lactic acid can activate the IL-23/IL-17 pathway, which is a canonical pro-inflammatory pathway [21]. However, lactic acid acts on tumor associated macrophages to polarize them toward an M2 state, augmenting tumor growth rates through a mechanism contingent on HIF-1α [33]. Therefore, lactic acid is an oncometabolite with important functions in cellular communication in the tumor microenvironment, which can reprogram immune cells, such as T-cells and macrophages, to become pathologic tumor-assisting agents. Moreover, the tumor microenvironment activates HIF-1α and AHR signaling to promote the metabolic reprogramming of immune cells and further modulate anti-tumor immunity [29, 30]. Thus, metabolic adaptation and the changes it imposes on the tumor microenvironment promote GBM survival and propagation by acting both on glioma and immune cells.
Recent studies have demonstrated that AHR plays an important role in the interplay between cancer metabolism and tumor-specific immunity. Tryptophan catabolism is increasingly recognized as a metabolic pathway that promotes tumorigenesis through its role in immune suppression [75]. The rate-limiting enzymes tryptophan-2,3-dioxygenase (TDO) and idoleamine-2,3-dioxygenase (IDO), expressed by tumor cells and antigen presenting cells, are thought to drive the increased catabolism of tryptophan in cancer [30, 76]. This metabolic pathway is up-regulated in several tumors including GBM, where it creates an immunosuppressive microenvironment through the depletion of the essential amino acid tryptophan and the generation of immunomodulatory tryptophan metabolites such as kynurenine (Kyn), which suppresses T cell function and induce apoptosis [77]. Importantly, Kyn is an agonist of AHR. Indeed, AHR activation by Kyn promotes Foxp3+ Treg differentiation, supporting earlier reports of a role of AHR in Tregs [9, 78]. Of note, AHR also promotes the differentiation of IL-10 producing Tr1 cells [7, 8], which together with Foxp3+ Tregs have been shown to contribute to tumor-associated immunosuppression [79]. In addition, AHR has also been linked to Th17 differentiation [79, 80]. However, detailed analyses suggest that AHR is more associated with non-pathogenic Th17 cells [80, 81]. Indeed, it has been shown that AHR drives the expression of the ectoenzyme CD39, which together with CD73 promotes the production of immunosuppressive adenosine. CD39-expressing Th17 cells with suppressive function have been associated with tumor immunosuppression.6
Almulla et al (2022) conducted a meta-analysis of the research into tryptophan and tryptophan catabolites (TRYCAT’s) in response to COVID-19. The pathway is activated due to oxidative stress and the activation of inflammatory cytokines:
Abstract
Background
Coronavirus disease 2019 (COVID-19) is accompanied by activated immune-inflammatory pathways and oxidative stress, which both induce indoleamine-2,3-dioxygenase (IDO), a key enzyme of the tryptophan (TRP) catabolite (TRYCAT) pathway. The aim of this study was to systematically review and meta-analyze the status of the TRYCAT pathway, including the levels of TRP and kynurenine (KYN) and the activity of IDO, as measured by the ratio of KYN/TRP.
Methods
This systematic review searched PubMed, Google Scholar, and Web of Sciences and included 14 articles that compared TRP and tryptophan catabolites (TRYCATs) in COVID-19 patients versus non-COVID-19 controls, as well as severe/critical versus mild/moderate COVID-19. The analysis was done on a total of 1269 people, including 794 COVID-19 patients and 475 controls.
Results
The results show a significant (p < 0.0001) increase in the KYN/TRP ratio (standardized mean difference, SMD = 1.099, 95% confidence interval, CI: 0.714; 1.484) and KYN (SMD = 1.123, 95% CI: 0.730; 1.516) and significantly lower TRP (SMD = − 1.002, 95%CI: − 1.738; − 0.266) in COVID-19 versus controls. The KYN/TRP ratio (SMD = 0.945, 95%CI: 0.629; 1.262) and KYN (SMD = 0.806, 95%CI: 0.462; 1.149) were also significantly (p < 0.0001) higher and TRP lower (SMD = − 0.909, 95% CI: − 1.569; − 0.249) in severe/critical versus mild/moderate COVID-19. No significant difference was detected in kynurenic acid (KA) and the KA/KYN ratio between COVID-19 patients and controls.
Conclusions
Our results indicate increased activity of the IDO enzyme in COVID-19 and severe/critical patients. The TRYCAT pathway is implicated in the pathophysiology and progression of COVID-19 and may signal a worsening outcome of the disease.
COVID-19 is characterized by activated immune-inflammatory pathways and, in some cases, hyperinflammation [5, 6]. Most importantly, during SARS-CoV-2 infection, the cytokine network is activated, with elevated levels of many pro-inflammatory cytokines such as interleukin (IL)-1β, IL-18, IL-6, tumor necrosis factor (TNF)-α, and interferon (IFN)-γ [7,8,9,10]. Mild COVID-19 may progress into SARS with pneumonia (and lowered oxygen saturation and lung lesions on chest computerized tomography scan), intravascular coagulation, multisystem failure, and death if these pro-inflammatory cytokines are overproduced during a cytokine storm [2, 7, 8]. Profound tissue damage, even extending to organ failure, may be the consequence of enduring increases in IFN-γ secretion [11]. COVID-19 is accompanied by increased production of reactive oxygen species (ROS) and ensuing oxidative damage, contributing to severe COVID-19 [12,13,14].
During infection, increased levels of IFN-γ, IL-1β, IL-6, and ROS may induce indoleamine-2,3-dioxygenase (IDO), which activates the catabolism of tryptophan (TRP), thereby lowering serum TRP and increasing tryptophan catabolites (TRYCATs), including kynurenine (KYN), 3-OH-kynurenine (3HK), kynurenic acid (KA), quinolinic acid (QA), and xanthurenic acid (XA) [15]. Activation of the TRYCAT pathway protects against hyperinflammation and microbial invasion by different processes including scavenging ROS, TRP starvation, and negative immunoregulatory effects [15, 16].
Furthermore, some TRYCATs, such as XA and KA, have antioxidant properties [17], whereas KYN, KA, XA, 3HK, and QA have negative immune regulatory effects, such as inhibiting IFN-γ production [16, 18]. Nonetheless, following overproduction of TRYCATs, several detrimental consequences may appear, including oxidative stress, immune activation, and neurotoxic effects [19,20,21,22,23,24,25].
In COVID-19, some authors reported increased activity of the TRYCAT pathway as indicated by lowered TRP and increased KYN levels and an increased KYN/TRP ratio [26,27,28], which reflects IDO activity [29]. Figure 1 shows the possible role of the TRYCAT pathway in COVID-19. Probably, the IDO enzyme, which is the first and rate-limiting enzyme of the TRYCAT pathway, is induced in COVID-19 by increased levels of IFN-γ, IL-1, IL-6, TNF-α, and ROS [15]. Moreover, stimulation of the aryl hydrocarbon receptor (AhR) by coronaviruses and IDO-induced KYN levels may cause the “systemic aryl hydrocarbon receptor activation syndrome” (SAAS), which aggravates hyperinflammation, hypercoagulation, and organ injuries [30]. It was hypothesized that TRYCAT pathway activation may worsen COVID-19 and probably decrease the patient's recovery potential [26, 31].
Summary of TRYCAT pathway in COVID-19. BMI body mass index, C cohort, T2DM type 2 diabetes mellitus, IFN-γ interferon-Gamma, IL-6 interleukin 6, IL-1β interleukin-1 beta, O&NS oxidative and nitrosative stress, O2 oxygen, CCTA chest computed tomography abnormalities, AhR aryl hydrocarbon receptor, IDO indoleamine 2,3 dioxygenase, TDO tryptophan 2,3-dioxygenase, KAT kynurenine aminotransferase, KMO kynurenine 3-monooxygenase, KYNU kynureninase, TRP tryptophan, KYN kynurenine, KA kynurenic acid, 3HK 3-hydroxykynurenine, AA anthranilic acid, XA xanthurenic acid, 3HA 3-hydroxyanthranilic acid, PA picolinic acid, QA quinolinic acid, NAD + nicotinamide adenine dinucleotide.
COVID-19 versus controls
The primary outcome variables KYN/TRP and (KYN + KA)/TRP ratio
The results of the systematic review on KYN/TRP in COVID-19 are shown in Table 1. We found that out of the ten included studies, the 95% CI for 7 (4 serum, 3 plasma) were entirely on the positive side of zero, while only one (plasma) study was totally on the negative side of zero. The two other studies showed 95% CI that overlapped with zero but with SMD values that were greater than zero. Figure 3 shows the forest plot of KYN/TRP in COVID-19 patients versus non-COVID-19 controls.
Forest plot with the results of a meta-analysis performed on the kynurenine/tryptophan (KYN/TRP) ratio in COVID-19 patients versus non-COVID-19 controls
Forest plot with the results of a meta-analysis performed on the kynurenine/tryptophan (KYN/TRP) ratio in severe/critical COVID-19 versus mild/moderate COVID-19
During infection, IDO activation and consequent increased TRYCATs but lowered TRP levels are key components of the innate immune response. First, the TRYCAT pathway has major intrinsic scavenging activities by neutralizing ROS [15]. Moreover, some TRYCATs have antioxidant properties on their own, as for example, 3-hydroxyanthranilic acid (3HA) and 3HK, which are more effective as radical scavengers than tocopherol, and XA, which has antioxidant activity comparable to that of butylated hydroxytoluene (BHT) [15, 17]. By protecting tissues from oxidative damage, KA has adequate antioxidant effects [61, 62]. Second, reduced TRP exerts anti-inflammatory (reduced T cell proliferation and activation, sensitization of apoptosis of activated T cells, and induction of the regulatory phenotype) and antimicrobial (inhibiting the growth of viruses, bacteria and parasites) effects through TRP starvation [63,64,65,66,67]. Third, TRYCATs such as KA, KYN, QA, and XA may have a negative immune-regulatory effect by decreasing IFN-γ production and/or increasing IL-10 production [15, 18]. In addition, KA has potent anti-inflammatory effects, while diminished KA levels may aggravate tissue damage and cell proliferation [68]. IFN-γ-induced stimulation of antigen-presenting cells upregulates the TRYCAT pathway and results in a counter-regulatory effect that preserves homeostasis [69]. Due to the fact that TRYCATs trigger apoptosis in Th-1, but not Th-2, cells, TRYCAT pathway activation may suppress Th-1 cells and promote Th-2 cell survival [70, 71]. As such, TRYCAT pathway activation results in a negative feedback loop to limit ROS production, hyperinflammation, and the Th-1 response [18, 70]. Fourth, some TRYCATs have neuroprotective effects including KA, anthranilic acid (AA) and XA. Thus, KA may inhibit N-methyl-D-aspartate (NMDA), kainate glutamate ionotropic, and amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, and reduce glutamate liberation through attenuating alpha 7 nicotinic acetylcholine receptors [16, 72]. XA inhibits vesicular glutamate transport (VGLUT), synaptic transmission via the NMDAR receptor, and excitatory postsynaptic potentials [73]. Furthermore, AA has neuroprotective effects by blocking the synthesis of neurotoxic TRYCATs such as picolinic acid (PA) and QA from 3HA [74].
Nevertheless, overproduction of some TRYCATs may cause detrimental effects on COVID-19. KA is implicated in deteriorating male COVID-19 patients through affecting the AhR, one of the master regulators of the immune-inflammatory response [75]. In addition, activation of AhR by TRYCATs, mainly KYN, affects immune resistance against viral infections and the airway basal cells of the lung epithelium, which are responsible for tissue repair [50, 76]. Most importantly, coronaviruses activate the same receptor through an IDO-independent mechanism while the IDO-AhR pathway in employed by viruses, bacteria, and parasites to establish infection [30]. Consequently, a positive feedback loop is established between increased TRYCATs levels due to IDO activation and stimulation of the AhR by TRYCATs and coronavirus [30]. Moreover, the AhR may enhance IDO transcription and regulate IDO activity [77]. These processes may result in the SAAS which may result in activated immune-inflammatory pathways (increased M1 cytokines), fibrosis (increased IL-22), thromboembolism (increased tissue factor and plasminogen activator inhibitor-1, consequent multiple organ injuries including brain injuries, and eventually death [30].
Role of TRYCAT pathway in comorbidities
Some TRYCATs have depressogenic, anxiogenic and neurotoxic effects, and TRYCATs like KYN are increased in neuropsychiatric illness including major depression, anxiety, and psychosis [15, 78]. Some TRYCATs exhibit pro-oxidant properties as evidenced by increased ROS, hydrogen peroxide, and superoxide production, and increased oxidative damage, including lipid peroxidation caused by 3HA, 3HK, and QA [19,20,21,22,23,24,25]. TRYCATs such as QA and XA and PA may have direct neurotoxic effects by activating hippocampal NMDAR and causing excitotoxicity with apoptosis and hippocampal shrinkage, thereby inducing neurocognitive impairments [79, 80]. Elevated XA levels may cause severe neuronal damage, apoptosis, mitochondrial dysfunctions, disrupt glutamate transmission, and impair presynaptic transmission caused by NMDAR stimulation [73]. Such effects may contribute to the development of neuropsychiatric disorders such as depression, anxiety and chronic fatigue due to COVID-19 [81]. Indeed, TRYCATs are confirmed to be associated with various mental disorders, including depression, and anxiety [15, 18], somatization and chronic fatigue syndrome [82], cognitive impairments [83], and psychosis [37]. Moreover, some TRYCATs, namely KYN, KA and 3HK are associated with musculoskeletal injuries due to their agonistic effects on the AhR [84,85,86,87]. Thus, increased TRYCAT levels could exacerbate the neuro-immune and neuro-oxidative toxicity caused by increased oxidative stress and M1 and Th-1 activation, resulting in comorbid affective disorders [81]. Therefore, it is safe to say that the accumulation of TRYCATs in SARS-CoV2-infected patients may play a role in the neuropsychiatric and cognitive syndromes of long or post-COVID syndrome [88].
Finally, it may be hypothesized that COVID-19-associated TRYCAT pathway activation may aggravate the existing disorders in this pathway in comorbid disorders (obesity, dementia, T2DM, hypertension and heart disease, stroke, chronic obstructive pulmonary disease (COPD) and chronic kidney disease) [1,2,3]. Indeed, in all those comorbid diseases, the IDO enzyme is activated as indicated by an increased KYN/TRP ratio [89,90,91,92,93,94,95]. By inference, when COVID-19 develops in people with those comorbid illnesses, an amplified TRYCAT response may occur, contributing to aggravated toxicity in addition to the consequences of inflammation and oxidative stress.7
A limited study of 150 patients, with 44 testing positive for COVID-19 by Santiago-Mujika et al (preprint, 2022) found that even a mild infection was associated with depleted tryptophan and increased levels of circulating neurotoxic metabolites via the KP.
Abstract
SARS-CoV-2 corona virus causes a multi-faceted and poorly defined clinical and pathological phenotype involving hyperinflammation, cytokine release. And long-term cognitive deficits, with an undefined neuropathological mechanism. Inflammation increases the activity of the kynurenine pathway, which is linked to neurodegenerative and psychiatric disorders. We sought to determine whether the kynurenine pathway is impacted in patients with mild COVID-19, leading to elevated neurotoxic metabolites in blood, and whether such changes are associated with pro-inflammatory cytokines. Serum samples were taken from 150 patients and analyzed by ELISA and ultra-high performance liquid chromatography (UHPLC). The data were analyzed using multiple linear regression models adjusted for age and sex. We found increased levels of kynurenine, quinolinic acid and 3-hydroxykynurenine in serum from patients with mild COVID-19, together with increased levels of IL-6, ICAM-1, VCAM-1 and neopterin. The levels of neurotoxic metabolites were significantly associated with key inflammatory cytokines including IL-6 and TNFα. The COVID-19 risk-factor hypertension was associated with the highest levels of neurotoxic metabolites in plasma. These neuroactive metabolites could be part of the pathological mechanisms underlying cognitive impairment during and post-COVID and should be explored as potential biomarkers for long-COVID symptoms.
Figure 1.Significantly higher levels IL-6 in patients with mild COVID-19 when compared to controls.
a) Significantly higher levels of IL-6 were found in patients with COVID-19 when compared to the negative controls (data adjusted for age and sex, ANOVA test F: 5.260, p=0.002 **). b) No differences were found in the level of TNF-α between the two groups (data adjusted for age and sex, ANOVA test F: 2.347, p= 0.075 ns). Graphs are represented by median with 95% of confidence interval (CI).
Figure 3.The kynurenine pathway is altered in patients with mild COVID-19, who present increased levels of neurotoxic metabolites.
Significantly increased levels of kynurenine (data adjusted for age and sex, ANOVA test F: 11.195, p<0.001 ***), 3-hydroxykynurenine (data adjusted for age and sex, ANOVA test F: 3.390, p=0.009 **), anthranilic acid (data adjusted for age and sex, ANOVA test F: 4.024, p=0.009 **), and quinolinic acid (data adjusted for age and sex, ANOVA test F: 8.492, p<0.001 ***) were found in patients with mild COVID-19 when compared to controls. When the ratio of the metabolites was analyzed, significantly increased levels of KYN/TRP (data adjusted for age and sex, ANOVA test F: 6.377, p<0.001 ***) and QUIN/TRP (data adjusted for age and sex, ANOVA test F: 5.837, p<0.001 ***), as well as QUIN/KYNA (data adjusted for age and sex, ANOVA test F: 2.847, p= 0.040 *) were found in patients with COVID-19. Graphs show the median with 95% of CI. Abbreviations: IDO, Indoleamine 2,3-dioxygenase; TDO, Tryptophan 2,3-dioxygenase; KATs, Kynurenine aminotransferase; KYNA, kynurenic acid; KMO, Kynurenine 3-monooxygenase; AA, anthranilic acid; 3-HK, 3-hydroxykynurenine; KYNU, Kynureninase; 3-HAA, 3-hydroxyanthranilic acid; 3HAO, 3-hydroxyanthranilate oxidase; ACMSD, Aminocarboxymuconate-semialdehyde decarboxylase, and NAD, Nicotinamide adenine dinucleotide.
Correlation between neurotoxic metabolites of the kynurenine pathway and pro-inflammatory cytokines
Previous research has shown the relationship between inflammation and the kynurenine pathway activity. Therefore, we subsequently investigated whether the levels of kynurenine metabolites and inflammatory cytokines were correlated. As shown in Figure 4, there was a positive correlation between the levels of TNF-α and KYN (Pearson correlation: 0.453; p=0.002**), 3-HK (Pearson correlation: 0.527; p<0.001***) and QUIN (Pearson correlation: 0.482; p<0.001***). Furthermore, a positive correlation between IL-6 and both 3-HK (Pearson correlation: 0.328; p=0.03*) and QUIN (data adjusted for age and sex, Pearson correlation: 0.418; p=0.005**) was also found. All data was corrected for age and sex.
Figure 4.There is correlation between inflammatory cytokines and metabolites of the kynurenine pathway in patients with mild COVID-19.
Patients with mild COVID-19 present a positive correlation between TNF-α and kynurenine (data adjusted for age and sex, Pearson R: 0.4512; p=0.0021 **), TNF-α and 3-hydroxykynurenine (data adjusted for age and sex, Pearson R: 0.4567; p=0.0018 **), TNF-α and quinolinic acid (data adjusted for age and sex, Pearson R: 0.4812; p=0.0009 ***), IL-6 and 3-hydroxykynurenine (data adjusted for age and sex, Pearson R: 0.3252, p=0.0312 *) and between IL-6 and quinolinic acid (data adjusted for age and sex, Pearson R: 0.4161, p=0.0050 **).
The correlation between IL-6, IFN-ɣ, and TNF-α and kynurenine metabolites supports the link between inflammation, SARS-CoV-2 and the kynurenine pathway. We found a positive correlation between TNF-α and three metabolites: 3-HK, KYN, and QUIN. TNF-α affects the kynurenine pathway in patients with schizophrenia, as a positive correlation between the levels of TNF-α and KYN has been reported (35). TNF-α may accelerate the formation of KYN from the catabolism of TRP (35). We did not observe differences in TNF-α levels between negative and positive SARS-CoV-2 patients, however, we did find a correlation between TNF-α and KYN levels, similar to what has been reported for schizophrenia (35). In this study, QUIN, 3-HK and KYN were significantly increased in patients with mild COVID-19. Both QUIN and 3-HK are neurotoxic metabolites and were positively correlated with TNF-α and IL-6 in the case of QUIN, and TNF-α in the case of 3-HK, it is possible that the inflammation observed in COVID-19 patients further contributes to the neuronal damage caused by the neurotoxic metabolites.
To summarize, we found an increase in neurotoxic metabolites of the kynurenine pathway in patients with mild COVID-19. Furthermore, the neurotoxic metabolites were correlated with inflammatory markers and vascular injury markers, such as TNF-α, IL-6, VCAM-1 and ICAM-1. We hypothesize that the activation of the neurotoxic branch of the kynurenine pathway might contribute to neurological, cognitive, and psychiatric symptoms experienced in COVID-19 and its aftermath. We suggest that these metabolites should be studied further for their potential as biomarkers of long COVID and as potential contributors to the disease mechanisms underlying long COVID.8
A second paper on tryptophan absorption, metabolism and long COVID, with more evidence to confirm the pathophysiology. Eroglu et al (2021):
Abstract
The global pandemic of COVID-19 has been lasting for more than one year and there is little known about the long-term health effects of the disease. Long-COVID is a new term that is used to describe the enduring symptoms of COVID-19 survivors. Huang et al. reported that fatigue, muscle weakness, sleep disturbances, anxiety, and depression were the most common complaints in COVID-19 survivors after 6 months of the infection. A recent meta-analysis showed that 80% of COVID-19 survivors have developed at least one long-term symptom and the most common five were fatigue, headache, attention deficit disorder, hair loss, and dyspnea. In this paper, we discuss the hypothesis that altered tryptophan absorption and metabolism could be the main contributor to the long-term symptoms in COVID-19 survivors.
A limited study of 33 patients testing positive for COVID-19 and 16 negative determined that tryptophan metabolism was the leading pathway affected:
Several studies have looked into changes in tryptophan metabolism in people infected with SARS-CoV-2 and found augmented activation of the KP. Thomas et al. [11] conducted a metabolomic study with the plasma of 33 participants who were positive for SARS-CoV-2 positive and 16 negative, and determined that tryptophan metabolism was the leading pathway affected by COVID-19. In those who were infected, they discovered lower levels of tryptophan, serotonin, and indolepyruvate, as well as higher levels of kynurenine, kynurenic acid, picolinic acid, and nicotinic acid, all of which were positively correlated with interleukin-6 levels. Despite the fact that the infection group was younger than the control group, this finding was significant because it demonstrated disturbed tryptophan metabolism in people infected with SARS-CoV-2.
This study found that the Kyn:Try ratio was also higher in those with severe lymphopenia:
In a separate study, Lionetto et al. [9] compared the serum Kyn:Try ratio—which reflects KP activation—in three groups: positive for SARS-CoV-2, negative for SARS-CoV-2 and admitted to the emergency department with illnesses other than COVID-19, and a healthy control group. The SARS-CoV-2-positive group had the highest Kyn:Try ratios. In the subgroup analysis, the SARS-CoV-2-positive participants with the most severe outcomes had the highest Kyn:Try ratios. The Kyn:Try ratio was also higher in those with severe lymphopenia, which is an ominous prognostic predictor in COVID-19, and in males, who are thought to be more vulnerable to infection[9]. Altered tryptophan metabolism in acute COVID-19 infection is also supported by several other studies.
KP and long-COVID symptoms
As discussed before, the most commonly seen long-term symptoms in people who have had COVID-19 are depression, fatigue, sleep disturbances, attention disorders, anxiety, muscle weakness, and dyspnea. When taken together, KP activation may also contribute these symptoms.
The “kynurenine shunt” refers to the increased degradation of tryptophan toward kynurenine and away from serotonin production [14]. Increased IDO activity has been related to depression in studies, owing to both serotonin depletion and neurotoxic effects of KP metabolites [15]. Fatigue, the most common long-term symptom in people who have had COVID-19, is divided into central and peripheral fatigue, with central fatigue causing complex weakness and making recovery difficult [16]. Several studies have revealed that increased metabolites of the KP in the brain trigger central fatigue and memory issues by inducing neurotoxicity [17], [18], [19], [20]. The long-term symptoms seen in COVID-19 infection could be related to the kynurenine shunt. Although there has been evidence of increased KP activation in acute COVID-19, there is no evidence in long-term COVID-19 survivors. Studies in this area can aid in our understanding of the pathophysiology underlying COVID-19’s long-term symptoms.9
However, a study of HIV patients by Routy et al (2015) found that the gut biome could be affected by viral protein activation of the KP via IDO upregulation, leading to a long term self sustaining feedback mechanism:
Abstract
Tryptophan degradation along the kynurenine pathway is associated with a wide variety of pathophysiological processes, of which tumor tolerance and immune dysfunction in several chronic viral infections including HIV are well known. The kynurenine pathway is at the crossroads of metabolism and immunity and plays an important role in inflammation while also playing an opposing role in the control of acute and chronic infections. In this review we have summarized findings from recent studies reporting modulation of tryptophan degrading the kynurenine pathway in the context of HIV infection. This immuno-metabolic pathway is modulated by three distinct inducible enzymes: indoleamine 2,3-dioxygenase 1 and 2 and tryptophan 2,3-dioxygenase. Increased expression of these enzymes by antigen-presenting cells leads to local or systemic tryptophan depletion, resulting in a mechanism of defense against certain microorganisms. Conversely, it can also lead to immunosuppression by antigen-specific T-cell exhaustion and recruitment of T regulatory cells. Recently, among these enzymes, indoleamine 2,3-dioxygenase 1 has been recognized to be an immune response checkpoint that plays an important role in HIV immune dysfunction, even in the context of antiretroviral therapy. In addition to the activation of the kynurenine pathway by HIV proteins Tat and Nef, the tryptophan-degrading bacteria present in the intestinal flora have been associated with dysfunction of gut mucosal CD4 Th17/Th22 cells, leading to microbial translocation and creating a systemic kynurenine pathway activation cycle. This self-sustaining feedback loop has deleterious effects on disease progression and on neurocognitive impairment in HIV-infected patients. Therapy designed to break the vicious cycle of induced tryptophan degradation is warranted to revert immune exhaustion in HIV-infected persons.10
This study by Sadlier et al (2022) of 20 patients with long COVID (PASC) also found a marked depletion of various metabolites, including serotonin:
Infection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) not only causes acute coronavirus 2019 (COVID-19) disease but also can lead to significant long-term effects that impact daily functioning and quality of life, termed postacute sequelae of COVID-19 (PASC).1The most frequently reported symptoms include fatigue, cognitive dysfunction, and memory issues, but multisystem involvement and significant disability are also common.2 Despite the rapid accumulation of knowledge on the acute phase of COVID-19, a limited number of studies are currently available that examine the pathophysiology of PASC, particularly in relation to fatigue. Mechanisms are thought to include the direct consequences of viral infection, severe systemic inflammation, oxidative stress, neuroinflammation, microvascular thrombosis, and neurodegeneration processes. We previously showed that levels of multiple cytokines, including TH2 cytokines such as interleukin (IL)-4, macrophage-derived chemokine (MDC), and thymic stromal lymphopoietin (TSLP), remained elevated for an extended period of time following SARS-CoV-2 infection.3 In this study, our aim was to identify additional potential novel mechanisms of disease, by investigating changes in metabolism that associate with PASC.
Serum was obtained from PASC patients (n = 20) at two time points (4–6 months following infection (T1) and 6–9 months following infection (T2)). All patients had been hospitalized for PCR-proven SARS-CoV-2 infection (median in-patient stay of 6.5 days, range 2 days to 25 days) during the first wave of the pandemic in Ireland (March–May 2020). The most common symptoms at follow-up clinics were fatigue and/or dyspnea.
Sphingosine-1-phosphate: “Although S1P is of importance in the entire human body, it is a major regulator of vascular and immune systems. In addition, it might be relevant in the skin. In the vascular system, S1P regulates angiogenesis, vascular stability, and permeability. In the immune system, it is now recognized as a major regulator of trafficking of T- and B-cells. S1P interaction with its receptor S1PR1 is needed for the egress of immune cells from the lymphoid organs (such as thymus and lymph nodes) into the lymphatic vessels. Inhibition of S1P receptors was shown to be critical for immunomodulation. S1P has also been shown to directly suppress TLR mediated immune response from T cells.[10]
A research team, led by a scientist at Weill Cornell Medical College, has discovered that red blood cells perform a second vital function: angiogenesis. Given its role in creating new blood vessels, scientists recognize S1P as vital to human health — and a player in some diseases, such as cancer. And although S1P is known to be blood borne, no one realized until this study that S1P is supplied by red blood cells to control blood vessel growth.”
“The levels of S1P (in a range of 5–40 μmol/L) are 5 to 10 times up-regulated in ovarian cancer patients' ascites. S1P at this physiological concentration stimulates migration and invasion of epithelial ovarian cancer cells but inhibits migration of normal ovarian surface epithelial cells.[11] Most (more than 90%) ovarian cancers arise from the epithelium of the ovary. Therefore, extracellular S1P could have an important role in cancer progression by promoting migration of epithelial ovarian cancer cells.”
“In addition, S1P modulates the proliferation of skin cells. This in particular applies to keratinocytes[13] while fibroblasts are not addressed in this way, apart from cell growth and differentiation While S1P suppresses epidermal proliferation as the glucocorticoids do, it differs from them in so far, as proliferation of dermal fibroblasts is not reduced. In fact, S1P even activates fibroblast-derived extracellular matrix protein production.”11
“Eicosanoid 12(S)-HETE upregulates endothelial cell alpha V beta 3 integrin expression and promotes tumor cell adhesion to vascular endothelium.”12
Distinct differences in circulating metabolite levels were evident in PASC patients compared to healthy volunteers (Figure 1). Of the 1086 metabolites quantified, levels of 253 metabolites or 266 metabolites were significantly different between patients and controls at T1 or T2, respectively (Tables S2 and S3). The metabolic profile of PASC patients observed at 6–9 months following SARS-CoV-2 infection (T2) was similar to the profile observed at 4–6 months (T1) following infection (Figure 1). Length of time stored in the freezer did not influence metabolite levels quantified in healthy volunteers or patient sera (Figure S1). Levels of multiple metabolites with immunomodulatory properties were elevated in PASC patients (examples include the sphingolipid sphingosine 1-phosphate (S1P) and the eicosanoid 12-HETE, Figure 2). In addition, significant disruption of central energy, fatty acid, and carbon metabolism was evident (examples include elevated mannose, glutamate, and succinate serum levels with reduced levels of the ketone body acetoacetate in PASC patients, Figure 2)
FIGURE 1 Principal component analysis of serum metabolite levels in healthy volunteers (Controls, n = 20) and PASC patients (n = 20) at 4–6 months (T1) and 6–9 months (T2) following hospital discharge
FIGURE 2 Violin plots illustrating the differences in specific serum metabolite levels in healthy volunteers (Controls, n = 20) and PASC patients (n = 20) at 4–6 months (T1) and 6–9 months (T2) following hospital discharge. Statistical significance was determined using one-way ANOVA and Tukey’s multiple comparison test
Of particular interest was the dysregulated metabolism of tryptophan, characterized by a decrease in serotonin production coupled with the accumulation of quinolinate in PASC patients (Figure 2). We have previously shown that tryptophan metabolism was heavily disrupted in COVID-19 patients during the acute phase of the infection, especially in those with the poorest outcomes.4Quinolinate is an excitotoxin that hinders neuronal function through multiple mechanisms including acting as an NMDA receptor agonist, and it indirectly drives accumulation of glutamate. In contrast, serotonin has essential modulatory effects on mood, anxiety, sleep, cognition, and memory. Recently, selective serotonin reuptake inhibitors (SSRIs) have been shown to have favorable results with respect to symptom resolution and hospitalizations in patients with COVID-19.
Our study contributes additional insights into underlying PASC mechanisms as we showed that infection-induced metabolic reprogramming and compensatory responses were long-lasting and did not substantially improve over the time course of the investigation (i.e., up to 9 months following the initial infection). These study findings identify novel mechanistic and diagnostic markers and potential therapeutic targets in PASC patients. These biomarkers should be included as integral components of future randomized controlled trials in order to better understand the pathobiology of SARS-CoV-2 infection and associated long-term sequelae.13
Many of the cases cited were from 2020, prior to elevated levels of population immunity and the rollout of experimental gene therapy agents under emergency use authorisations.
Post transfection exposure to or expression of spike protein for months due to persistence in the lymph node germinal centres, on monocytes or due to reverse transcription in various tissues may offer an explanation for the majority of currently reported cases of LC, as would repeated, high viral load infections due to immunosuppression and especially CD4+ T-cell depletion.
The resultant highly elevated sytemic levels of spike proteins trigger ROS generation14 by binding to ACE2 and elevate levels of inflammatory cytokines such as Il-6 & TNFa, both of which mediate tryptophan depletion and the KP with a disrupted, inflammatory gut biome feedback loop, as discussed earlier.
Indeed it is highly likely that transfection is responsible for the vast majority of so called “long COVID” cases currently being reported. These should perhaps be more accurately referred to as “vaccination-related adverse events”.
Therapeutics and Breaking the Cycle
The allopathic approach recommended so far included supplementation with tryptophan and the administration of selective serotonin reuptake inhibitors (SSRIs) to try to correct effects of the serotonin deficit caused by the KP.
But although it may help in the short term it is more akin to treating the symptoms rather than any of the many possible causes of long COVID, and there is no reason that multiple factors may be at work at the same time (ie pleiotropically), for example mitochondrial damage15, NAD+ deficiency, microinfarctions, fibrosis, autoimmune disorders16, linoleic acid deficit17, vagus nerve disruption18 or GP120 neurotoxicity.19
Additionally, long term taking of SSRI’s has been linked to increased tendencies towards suicide:
Abstract
Objective: The authors explored the relationship between the initiation of therapy with selective serotonin reuptake inhibitor (SSRI) antidepressants and completed suicide in older patients.
Method: The authors linked population-based coroner's records with patient-level prescription data, physician billing claims, and hospitalization data for more than 1.2 million Ontario residents 66 years of age and older from 1992 to 2000. For each suicide case, four closely matched comparison subjects were selected using propensity score methods. The authors determined the odds ratio for suicide with SSRIs versus other antidepressant treatment, calculated at discrete monthly intervals from the start of treatment.
Results: Of 1,329 suicide cases, 1,138 (86%) were each fully matched to four comparison subjects using propensity scores. During the first month of therapy, SSRI antidepressants were associated with a nearly fivefold higher risk of completed suicide than other antidepressants (adjusted odds ratio: 4.8, 95% confidence interval=1.9-12.2). The risk was independent of a recent diagnosis of depression or the receipt of psychiatric care, and suicides of a violent nature were distinctly more common during SSRI therapy. Numerous sensitivity analyses revealed consistent results. No disproportionate suicide risk was seen during the second and subsequent months of treatment with SSRI antidepressants, and the absolute risk of suicide with all antidepressants was low.
Conclusions: Initiation of SSRI therapy is associated with an increased risk of suicide during the first month of therapy compared with other antidepressants. The absolute risk is low, suggesting that an idiosyncratic response to these agents may provoke suicide in a vulnerable subgroup of patients.20
A more holistic approach should be considered to address the root cause for induction and sustenance of the kynurenine pathway, for example:
To disrupt the expression of spike protein due to transfection or persistent viral infection by taking antivirals such as ivermectin, berberine21, Artemisia annua22 or a range of other therapeutics with broad spectrum antioxidant and anti-inflammatory properties to disrupt the KP.23 Echinacea also has excellent T-cell stimulatory, antiviral and cytokine inhibiting effects.24
Enzymically degrading any legacy spike protein is challenging due to its resistant, prion-like properties. Resveratrol or proteolytic enzymes such a serrapeptase or nattokinase25 may be of therapeutic value.
L-Tryptophan supplementation to balance moderation of the kynurenine pathway. It is often sold to body builders to bulk-up muscle mass. Muscle wastage can be another symptom of LC due to the Warburg Effect.
A probiotic diet to improve the gut biome to a less inflammatory state so that tryptophan depletion and the KP is moderated, to break the cycle, improve immunity and help facilitate recovery.
Kechagia et al (2013) reviewed the health benefits of probiotic foods:
Abstract
Probiotic bacteria have become increasingly popular during the last two decades as a result of the continuously expanding scientific evidence pointing to their beneficial effects on human health. As a result they have been applied as various products with the food industry having been very active in studying and promoting them. Within this market the probiotics have been incorporated in various products, mainly fermented dairy foods. In light of this ongoing trend and despite the strong scientific evidence associating these microorganisms to various health benefits, further research is needed in order to establish them and evaluate their safety as well as their nutritional aspects. The purpose of this paper is to review the current documentation on the concept and the possible beneficial properties of probiotic bacteria in the literature, focusing on those available in food.
1. Introduction
The association of probiotics with well-being has a long history. More than a century has passed since Tissier observed that gut microbiota from healthy breast fed infants were dominated by rods with a bifid shape (bifidobacteria) which were absent from formula fed infants suffering from diarrhoea, establishing the concept that they played a role in maintaining health. Since then a series of studies have supported this association but they were originally poorly designed and controlled and faced practical challenges such as strain specificity of properties and the slow growth of probiotics in substrates other than human milk. By time, they have successfully evolved with the more recent ones accumulating more substantial evidence that probiotic bacteria can contribute to human health. These data have coincided with the increasing consumer awareness about the relationship between health and nutrition creating a supporting environment for the development of the functional food concept introduced to describe foods or food ingredients exhibiting beneficial effects on the consumers' health beyond their nutritive value. The functional food market is expanding, especially in Japan—its birthplace—with further growth prospects in Europe and the United States and in most countries the largest share of its products is held by probiotics [1, 2]. The reported beneficial effects of probiotic consumption include improvement of intestinal health, amelioration of symptoms of lactose intolerance, and reduction of the risk of various other diseases, and several well-characterized strains of Lactobacilli and Bifidobacteria are available for human use [3, 4]. Nevertheless, despite the promising evidence, the role of probiotics in human health as well as the safety of their application should be further investigated as the current knowledge of the characteristics that are necessary for their functionality in the gut is not complete.
5. Mechanisms of Probiotic Activity
Probiotics have various mechanisms of action although the exact manner in which they exert their effects is still not fully elucidated. These range from bacteriocin and short chain fatty acid production, lowering of gut pH, and nutrient competition to stimulation of mucosal barrier function and immunomodulation. The latter in particular has been the subject of numerous studies and there is considerable evidence that probiotics influence several aspects of the acquired and innate immune response by inducing phagocytosis and IgA secretion, modifying T-cell responses, enhancing Th1 responses, and attenuating Th2 responses [20–22].
7. Health Benefits of Probiotics
There is increasing evidence in favour of the claims of beneficial effects attributed to probiotics, including improvement of intestinal health, enhancement of the immune response, reduction of serum cholesterol, and cancer prevention. These health properties are strain specific and are impacted by the various mechanisms mentioned above. While some of the health benefits are well documented others require additional studies in order to be established. In fact, there is substantial evidence to support probiotic use in the treatment of acute diarrhoeal diseases, prevention of antibiotic-associated diarrhoea, and improvement of lactose metabolism, but there is insufficient evidence to recommend them for use in other clinical conditions.
12. Other Health Benefits
The list of health benefits mediated by probiotics is not limited to the ones mentioned so far and includes a range of promising effects that require however further human studies in order to be substantiated. There is evidence that probiotic bacteria are dietary components that may play a role in decreasing cancer incidence. The exact mechanisms are under investigation, but studies have demonstrated that certain members of Lactobacillus and Bifidobacterium spp. decrease the levels of carcinogenetic enzymes produced by colonic flora through normalization of intestinal permeability and microflora balance as well as production of antimutagenic organic acids and enhancement of the host's immune system [55, 56]. Furthermore, evidence suggests that food products containing probiotic bacteria could possibly contribute to coronary heart disease prevention by reducing serum cholesterol levels as well as to blood pressure control. Proposed mechanisms include interference with cholesterol absorption from the gut, direct cholesterol assimilation, and production of end fermentation products that affect the systemic levels of blood lipids and mediate an antihypertensive effect. Nevertheless, these probiotic effects are still a matter of debate as further research is needed in long-term human studies [57]. Last but not least, probiotic strains administered in dairy products have shown to improve the therapeutic outcome in women with bacterial vaginosis, most probably by supporting the normal vaginal lactobacilli microbiota.26
Probiotic bacteria and foods:
7 types of probiotic bacteria:
Lactobacillus acidophilus
Lactobacillus bulgarius
Lactobacillus reuteri
Streptococcus thermophilus
Saccharomyces boulardii
Bifidobacterium bifidum
Bacillus subtilis
The best probiotics naturally have these active cultures. It’s important they’re live and active cultures to benefit your gut the most, so if you do choose to take probiotic supplements, make sure they have these.
17 Top Probiotic Foods
Where can you find probiotics? Here are the top 17 foods in which you can find this beneficial bacteria.
1. Kefir
Similar to yogurt, this fermented dairy product is a unique combination of milk and fermented kefir grains. It has been consumed for well over 3,000 years; the term originated in Russia and Turkey and means “feeling good.” It has a slightly acidic and tart flavor and contains anywhere from 10 to 34 strains of probiotics.
It is similar to yogurt, but because it is fermented with yeast and more bacteria, the final product is higher in probiotics and lower in lactose, making it a suitable choice for many who are lactose-intolerant.
2. Sauerkraut
Made from fermented cabbage and other probiotic vegetables, sauerkraut is not diverse in probiotics but is high in organic acids (what gives food its sour taste) that support the growth of good bacteria.
Sauerkraut is extremely popular in Germany today. It is high in vitamin C and digestive enzymes. It’s also a good source of natural lactic acid bacteria, such as lactobacillus.
3. Kombucha
Kombucha is an effervescent fermentation of black tea that is started by using a SCOBY, also known as a symbiotic colony of bacteria and yeast. Kombucha has been around for over 2,000 years, originating around Japan. Many claims have been made about kombucha, but its primary health benefits include digestive support, increased energy and liver detoxification.
4. Coconut Kefir
Made by fermenting the juice of young coconuts with kefir grains, this option has some of the same probiotics as the traditional variety but is typically not as high in probiotics. Still, it has several strains that are beneficial for your health.
Coconut kefir has a great flavor, and you can add a bit of stevia, water and lime juice to make a great-tasting, refreshing drink.
5. Natto
A popular dish in Japan consisting of fermented soybeans, natto contains the extremely powerful probiotic Bacillus subtilis, which has been proven to bolster your immune system, support cardiovascular health and enhance digestion of vitamin K2.
Natto also contains a powerful anti-inflammatory enzyme called nattokinase that has been proven to prevent blood clotting and is loaded with protein, securing it a top slot in the list of probiotic foods.
6. Yogurt
Possibly the most popular probiotic food is live cultured probiotic yogurt or Greek yogurt made from the milk of cows, goats or sheep. Yogurt, in most cases, can rank at the top of probiotic foods if it comes from grass-fed animals and has not been pasteurized.
The problem is there is a large variation on the quality of yogurts on the market today. When buying yogurt, look for organic, grass-fed varieties that are made from goat’s or sheep’s milk.
7. Kvass
This powerful ingredient has been a common fermented beverage in Eastern Europe since ancient times. It was traditionally made by fermenting rye or barley, but in more recent years has been created using probiotic fruits and beets along with other root vegetables like carrots.
Kvass uses Lactobacilli probiotics and is known for its blood and liver-cleansing properties along with its mild sour flavor.
8. Raw Cheese
Goat’s milk, sheep’s milk and A2 cow’s soft cheeses are particularly high in probiotics, including thermophillus, bifudus, bulgaricus and acidophilus. Always buy raw and unpasteurized cheeses if you want to receive any probiotics, as pasteurized and processed varieties are lacking in beneficial bacteria.
9. Apple cider vinegar
Is apple cider vinegar a good source of probiotics? In addition to controlling blood pressure, reducing cholesterol levels, improving insulin sensitivity and even enhancing weight loss, apple cider vinegar can also help ramp up probiotic intake as well. Drink a small bit each day or use it as a salad dressing to maximize your results.
10. Salted gherkin pickles
These fermented tasty treats are also a little recognized probiotics source. When shopping for pickles, be sure to choose a smaller food manufacturer that uses organic products. If you can find a local maker, you’ll be getting some of the best probiotics for your health.
11. Brine-cured olives
Olives that are brine-cured are an excellent source of probiotics. Like with salted gherkin pickles, be sure to select a product that is organic first. Next, be certain that your olives aren’t made from a huge manufacturer and try to select a smaller company that advertises probiotics.
Also make sure that your olives don’t contain sodium benzoate, a food additive that can negate many of the health-promoting properties of this probiotic power-food.
12. Tempeh
Hailing from Indonesia, this fermented soybean product is another awesome food that provides probiotics. Tempeh is created by adding a tempeh starter to soybeans. The product is then left to sit for a day or two, which results in a cake-like product.
You can eat tempeh raw or by boiling it and eating it with miso. It can also be used as a substitute for meat in a stir fry meal and can be baked, grilled, marinated or sautéed.
13. Miso
Miso is a traditional Japanese spice found in many of their traditional foods. If you’ve ever been to a Japanese restaurant, you may have seen their miso soup. Not only that, but it is also one of the mainstays of Japanese medicine and is commonly used in macrobiotic cooking as a digestive regulator.
It is created by fermenting soybean, barley or brown rice with koji. Koji is a fungus, and the fermentation process takes anywhere from a few days to a few years to complete.
Miso soup is famous throughout the world, and it’s very easy to prepare. Simply dissolve a tablespoonful of miso in a pot of water filled with seaweed and other ingredients of your choice. Miso can also be spread on crackers, used in place of butter or added to marinades and stir-fries for an added dose of flavor.
14. Traditional Buttermilk
Traditional buttermilk, also sometimes called cultured buttermilk, is a fermented drink that is made from the liquid that is left over after churning butter. It’s considered one of the top probiotic Indian foods and is also commonly consumed in countries such as Nepal and Pakistan as well.
Keep in mind that most types of buttermilk found at supermarkets do not contain probiotics. Instead, look for varieties that contain live cultures to boost the benefits of your buttermilk.
15. Water Kefir
Water kefir is is made by adding grains to sugar water, resulting in a fermented, fizzy beverage that is jam-packed with probiotics.
The water kind is one of the top natural vegan probiotic foods that can be enjoyed as part of a healthy plant-based diet. It’s also thinner than the regular version and can be flavored using a variety of herbs, fruits and spices to create your own customized concoction.
16. Raw Milk
Raw cow’s milk, goat’s milk, sheep’s milk and A2 aged cheeses are particularly high in probiotics. Just remember, all pasteurized dairy is devoid of healthy bacteria, so to get the probiotics, you need to stick to only high-quality, raw dairy that hasn’t been pasteurized.
17. Kimchi
Kimchi is a cousin to sauerkraut and is the Korean take on cultured veggies. It’s created by mixing a main ingredient, such as Chinese cabbage, with a number of other foods and spices, like red pepper flakes, radishes, carrots, garlic, ginger, onion, sea salt and fish sauce.
The mixture is then left aside to ferment for three to 14 days, resulting in a flavor-filled, probiotic-packed ingredient.27
Thank you for reading DoorlessCarp’s Scientific Literature Review. This post is public so feel free to share it.
Routy JP, Routy B, Graziani GM, Mehraj V. The Kynurenine Pathway Is a Double-Edged Sword in Immune-Privileged Sites and in Cancer: Implications for Immunotherapy. Int J Tryptophan Res. 2016 Oct 12;9:67-77. doi: 10.4137/IJTR.S38355. PMID: 27773992; PMCID: PMC5063567.
Cai, C., Zhang, X., Liu, Y. et al.Gut microbiota imbalance in colorectal cancer patients, the risk factor of COVID-19 mortality. Gut Pathog 13, 70 (2021). https://doi.org/10.1186/s13099-021-00466-w
Luiz Gonzaga Francisco de Assis Barros D'Elia Zanella. Pathophysiology of COVID-19: SARS-CoV-2 mimics neoplastic cells and The Long COVID-19 Syndrome can be a Warburg Effect and ACE-2 internalization consequence, 11 May 2022, PREPRINT (Version 1) available at Research Square [https://doi.org/10.21203/rs.3.rs-1643416/v1]
Almulla, A.F., Supasitthumrong, T., Tunvirachaisakul, C. et al.The tryptophan catabolite or kynurenine pathway in COVID-19 and critical COVID-19: a systematic review and meta-analysis. BMC Infect Dis 22, 615 (2022). https://doi.org/10.1186/s12879-022-07582-1
Estibaliz Santiago-Mujika, Kevin Heinrich, Sonia George, Colt D Capan, Cameron Forton, Zachary Madaj, Amanda R Burmeister, Matthew Sims, J Andrew Pospisilik, Patrik Brundin, Stewart F Graham, Lena Brundin, Increased levels of circulating neurotoxic metabolites in patients with mild Covid19. (2022).
Routy JP, Mehraj V, Vyboh K, Cao W, Kema I, Jenabian MA. Clinical Relevance of Kynurenine Pathway in HIV/AIDS: An Immune Checkpoint at the Crossroads of Metabolism and Inflammation. AIDS Rev. 2015 Apr-Jun;17(2):96-106. PMID: 26035167.
Sadlier, C., Albrich, W.C., Neogi, U., Lunjani, N., Horgan, M., O’Toole, P.W. and O’Mahony, L. (2022), Metabolic rewiring and serotonin depletion in patients with postacute sequelae of COVID-19. Allergy, 77: 1623-1625. https://doi.org/10.1111/all.15253
Juurlink DN, Mamdani MM, Kopp A, Redelmeier DA. The risk of suicide with selective serotonin reuptake inhibitors in the elderly. Am J Psychiatry. 2006 May;163(5):813-21. doi: 10.1176/ajp.2006.163.5.813. PMID: 16648321.
Woooow!!!! Amaaaaaazing work!!!! Feel like I have just witnessed a BIG BANG Explosion of knowledge.
DoorlessCarp since you mentioned Astaxanthin and a lot of similar vitamins to what Patrick Theut recommends as part of his "Reverse CAC score & Heart Disease" I am sharing the link (pls see below) for Longecity.org forum on how to reverse arterial plaque and also an Abstract from Patrick Theut's interview with Ivor Cummins, your perusal.
Feel Free to delete my post if there is anything you feel uncomfortable about it. The only reason I am sharing it is because I think it might be of value to you and your readers. And it is a great protocol as well as Interview. The whole approach to reversing arterial plaque is awesome and I have not heard the better one anywhere.
HDL & LDL will kill & clean out macrophage foam cells with high serum levels of the following:
• LDL
• HDL
• Free T3 between 3.3 & 5.0 pg/ml
• K1, MK-4, & MK-7 koncentrated k supplies all these
• CoQ10
• Vit D3 vit-D, 25-OH between 60 &70
• Vit E
• Vit C Patrick takes 6 g/d, but realizes that is too high for most people
• Magnesium
• Selenium optimum is 200 ug/day (total including food)
• Omega 3's with fat [I take DHA & EPA extracts and avoid fish oil]
• Do not take Calcium or Arginine
• Take Lysine & Citrulline
Abstract from Patrick Theut's interview with Ivor Cummins plus [Editorial comments]
• Heart disease reversal/prevention requires high serum levels of vitamin K1 and vitamin MK-7 .
• Vitamin K: There is one K1 and 13 [misspoken] K2's. [Edit: The K2's are MK-4 thru MK-13, where the number indicates the length of a molecule chain. So there are 10 of them.]
• K1 is made by plants. It's high in broccoli & Kale.
• K2's come from fermented food products or fermentation by gut bacteria. Carbs feed the wrong type of gut bacteria. Carbs can kill K2 production in your gut. A low carb ketogenic diet is essential for the gut to make K2s. [Edit: Its best to supplement K2's instead of worrying about gut bacteria making them.]
Need for K1, vit-C, and "large fluffy" LDL
• Macrophages inside your cell wall turn into foam cells that cause constant inflammation and the build up of plaque.
• Initially, monocytes patrolling the inside of arteries pull damaged (gluconated or oxidized) lipids and bacteria out of the blood, then become macrophages inside the cell wall.
• Once macrophages job is done they need to be killed, else they become "foam cells", bloated with droplets of cholesterol.
• LDL carries K1 & vit-C. If it has enough K1 and Vit-C it will kill the macrophages that eat it. If there is not enough of either K1 or Vit-C, the macrophages will gorge on LDL and become foam cells.
• This explains why a vegetarian diet high enough in K1 and vit-C could prevent athersclerosis.
• Small LDL cannot carry enough K1, so small LDL will always feed foam cells. You need large fluffy LDL and enough K1 & Vit-C in order to stay healthy.
Need for vit-D & free-T3
• You've got macrophages and foam cells growing and you've got to kill them.
• If you have enough vit-D and enough free T3 (thyroid hormone), there will be proteins on the surface of macrophages that start spinning. This attracts HDL.
Need for MK-7, CoQ10, Micro-RNA, and "large fluffy" HDL
• MK-7 on the HDL locks onto the spinning protein on macrophages or foam cells and locks it down.
• The COQ10 on HDL then binds tightly to the macrophage. Then Micro-RNA from the HDL enters the macrophage (or foam cell) and kills it.
• The HDL then sucks out the fat & cholesterol and when its all bloated it returns to the liver.
• Small HDL cannot carry enough MK7, CoQ10, & Micro-RNA, so it cannot kill foam cells.
• The difference in surface area between small & large HDL is 900%.
• In addition, small HDL cannot orient MK7 & CoQ10 properly to lock onto a macrophage.
Need for magnesium
• A little-known carrier protein is attracted into the artery wall by spinning proteins on macrophages.
• If there is enough magnesium to loosen calcium from muscle cells in the arterial wall, this carrier protein can grab onto the calcium and remove it.
Diet
• Inadequate diet will make LDL & HDL too small
• Will not provide enough K1, vit-C, vit-D, free-T3, MK7, CoQ10, and magnesium.
• High dietary levels create more K1 & vit-C on in LDL
• High dietary levels create a better coating of MK-7, CoQ-10, and Micro-RNA on HDL
• High dietary levels of vit-D and free-T3 create the spinning molecules on macrophages
HDL & LDL will kill & clean out macrophage foam cells with high serum levels of the following:
• LDL
• HDL
• Free T3 between 3.3 & 5.0 pg/ml
• K1, MK-4, & MK-7 koncentrated k supplies all these
• CoQ10
• Vit D3 vit-D, 25-OH between 60 &70
• Vit E
• Vit C Patrick takes 6 g/d, but realizes that is too high for most people
• Magnesium
• Selenium optimum is 200 ug/day (total including food)
• Omega 3's with fat [I take DHA & EPA extracts and avoid fish oil]
• Do not take Calcium or Arginine
• Take Lysine & Citrulline
• One thing to add, since not many would take the time to watch long videos, is that Patrick since almost 2 decades has sieved anything he could find in the available scientific literature (his product page is a testimony to that in respect to K-vitamins only), and is propably the only individual who lab-tested himself the most.
•
• For example, by repeatedly testing undercarboxylated osteocalcin as a proxi for vitamin K2, he found that no high doses of K2 vitamins can be too much, to bring it down to healthier levels. So he know's his own metabolism very well. And many of his recommentations originate from that.
•
• Doing a lot of lab-testing myself (not even nearly as much), I want to point out that there are immense differences in bio-chemical individuality. For example my selenium only came in the optimal range with 370 mcg/d of selenium (incl. food).
• He thinks a carrot a day gives enough preforemed vitamin A. And the liver know's by itself how much is actually needed. In his later video he admits that there are of course genetic differences. I also found carrots not enough, but the need for high doses of preformed vitamin A to raise serum levels to optimal.
• My biome showed I've complete lack in vitamin K1 to K2 metabolizing bacteria, despite being low carb since a decade.
• Only Mg-sulfate IVs, beside supplementing up to 2.5 g of oral elemental Mg a day, finally started to improve my severe Mg-deficiency.
• My rhinitis eats up so much vitamin C for its antihistamine properties, I actually supplemented 24 g/d, and would still do better with more of that stuff.
• So take what it takes, acertained with frequent lab-testing.
•
• Despite obviously being deficient in freeT3 and Mg, I too could halt the mercyless growth in calcification. We are all deferent.
started taking C60oo in early August, 2012, and it has reduced the depth of the wrinkles on my face, as seen in the photos on my profile.
I also noticed fading of two deep scars after Turnbuckle noted that he saw scars fading, likely due to C60oo's potent antioxidant effect improving the health of skin's elastin and collagen.
I assume that the external anti-aging improvements that I've experienced are accompanied by internal improvements.
However, while C60oo appears to reduce the potential for Alzheimer's, cancers, liver inflammation and other potential problems that would limit lifespan, it does not appear to affect cardiovascular issues - blood lipids, etc...
So, with finding out that I have some plaque, I began researching the potential to reverse arterial plaque and improve overall cardiovascular function with a goal of living far past 100 years of age in good health.
I then found a study that showed that high-dose vitamin K2 (MK4) reversed arterial plaque in rats ~37% in just six weeks.




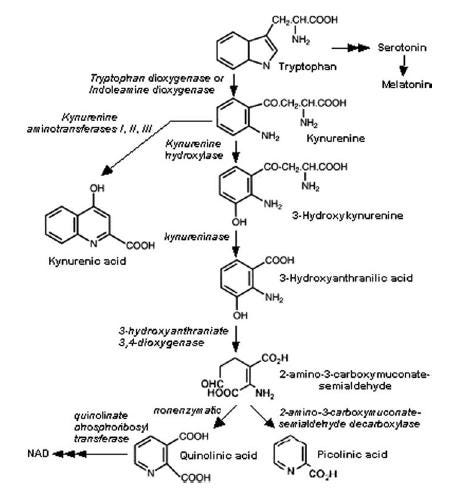

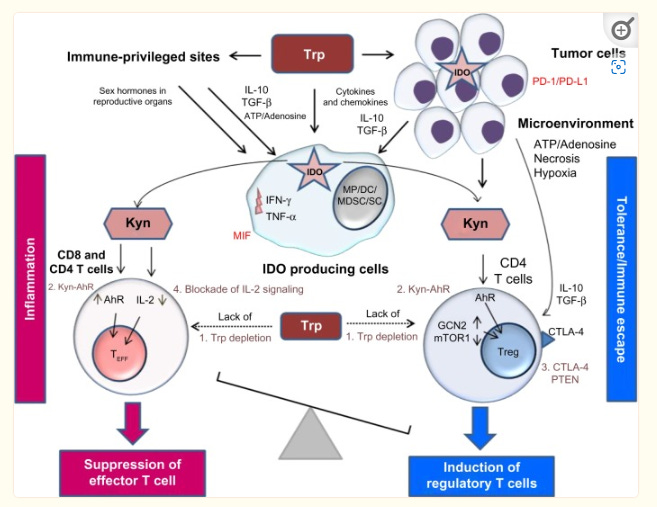

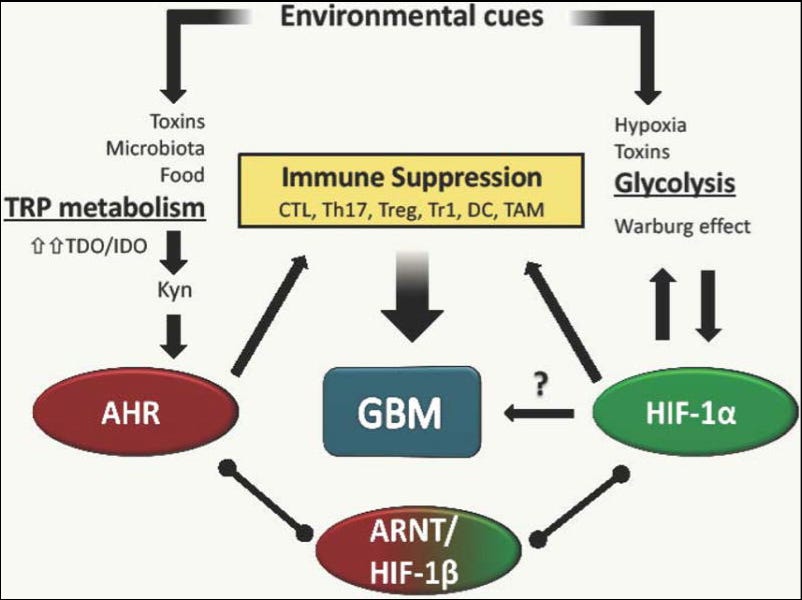



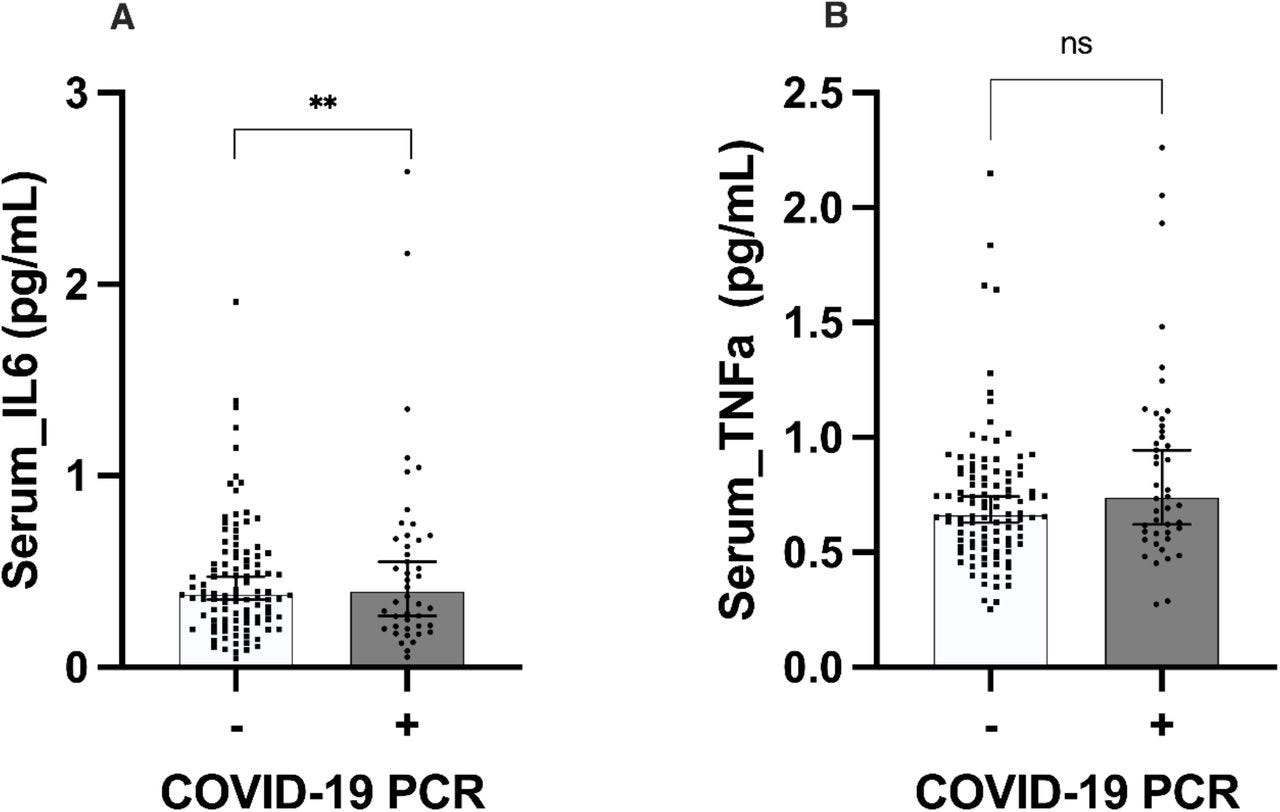
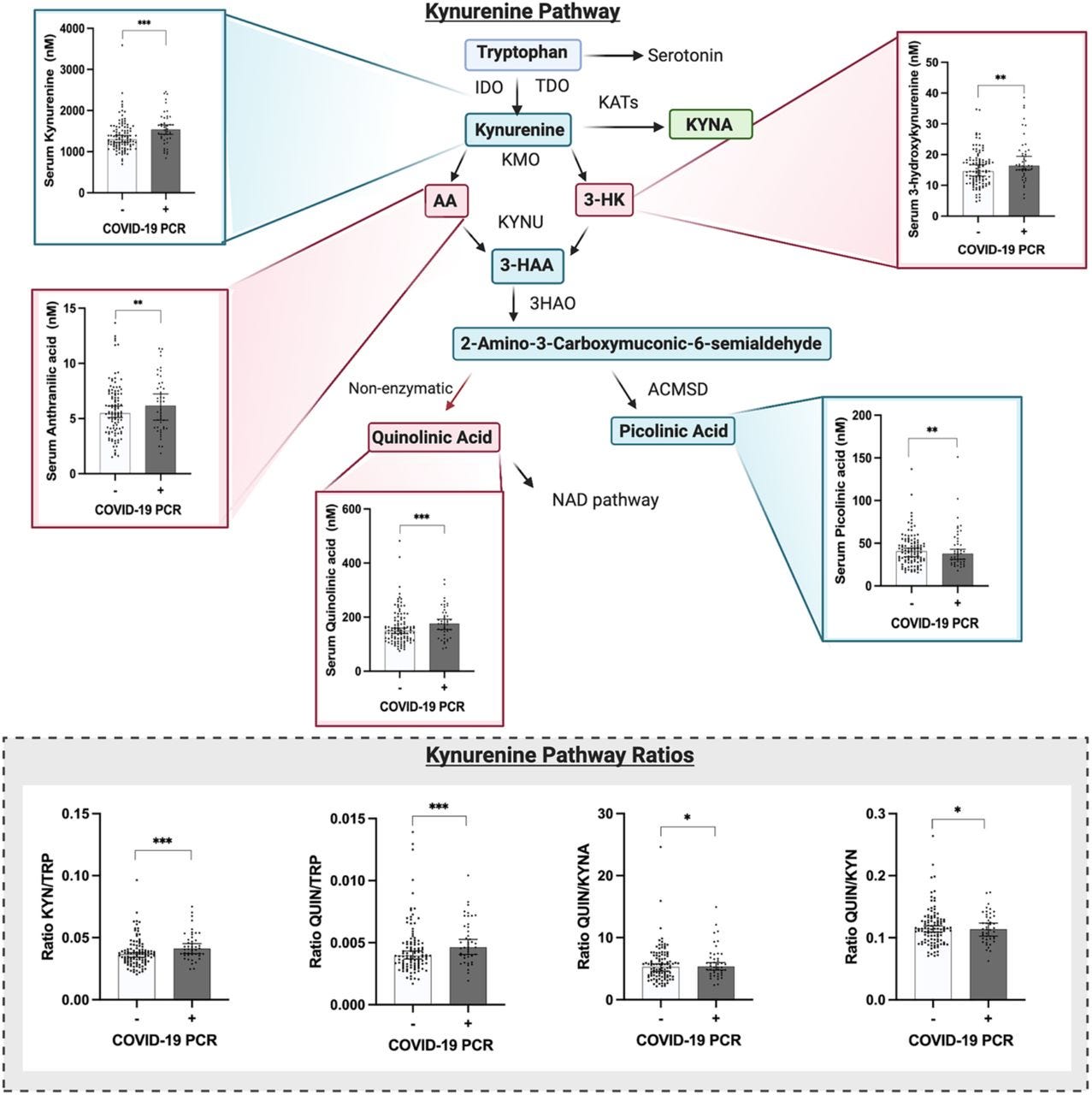
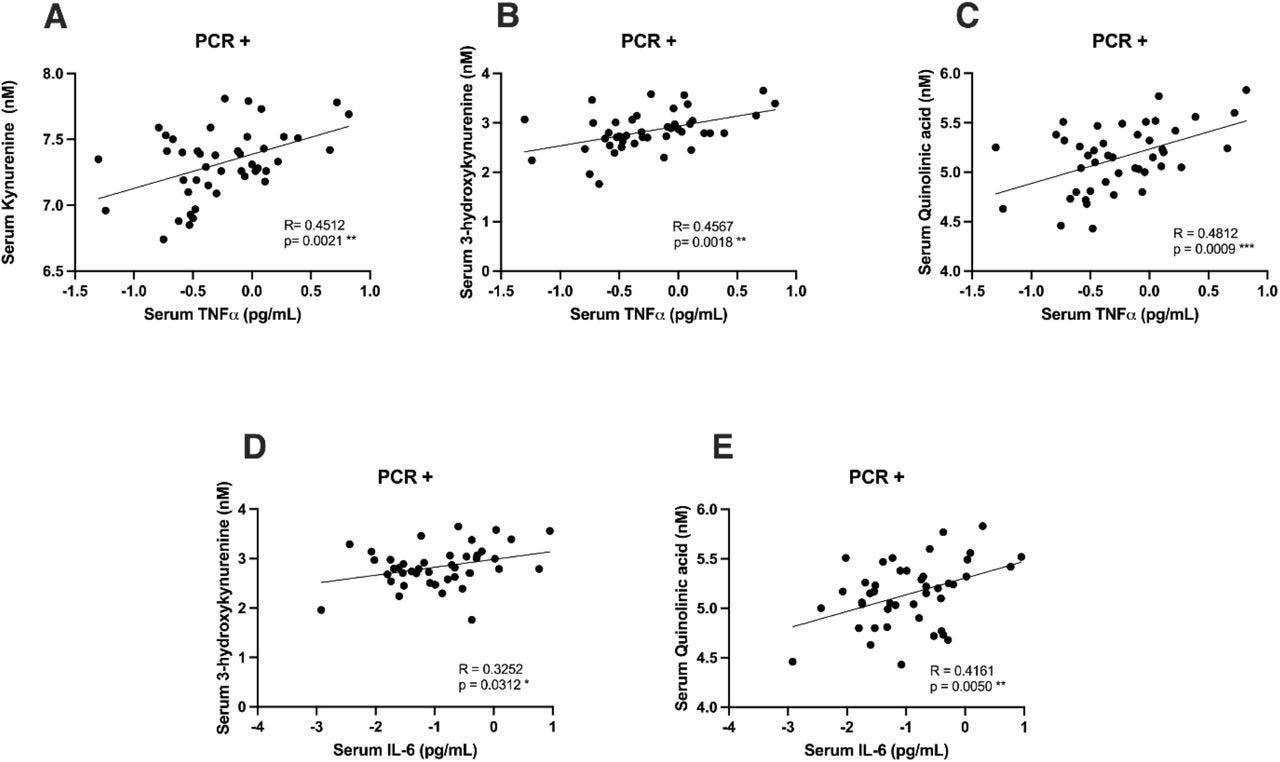




Woooow!!!! Amaaaaaazing work!!!! Feel like I have just witnessed a BIG BANG Explosion of knowledge.
DoorlessCarp since you mentioned Astaxanthin and a lot of similar vitamins to what Patrick Theut recommends as part of his "Reverse CAC score & Heart Disease" I am sharing the link (pls see below) for Longecity.org forum on how to reverse arterial plaque and also an Abstract from Patrick Theut's interview with Ivor Cummins, your perusal.
Feel Free to delete my post if there is anything you feel uncomfortable about it. The only reason I am sharing it is because I think it might be of value to you and your readers. And it is a great protocol as well as Interview. The whole approach to reversing arterial plaque is awesome and I have not heard the better one anywhere.
https://www.longecity.org/forum/topic/65884-reversing-arterial-plaque/page-1
https://www.longecity.org/forum/topic/111100-patrick-theut-reverse-cac-score-heart-disease/?ref=tfrecipes
Patrick Theut "Reverse CAC score & Heart Disease"
HDL & LDL will kill & clean out macrophage foam cells with high serum levels of the following:
• LDL
• HDL
• Free T3 between 3.3 & 5.0 pg/ml
• K1, MK-4, & MK-7 koncentrated k supplies all these
• CoQ10
• Vit D3 vit-D, 25-OH between 60 &70
• Vit E
• Vit C Patrick takes 6 g/d, but realizes that is too high for most people
• Magnesium
• Selenium optimum is 200 ug/day (total including food)
• Omega 3's with fat [I take DHA & EPA extracts and avoid fish oil]
• Do not take Calcium or Arginine
• Take Lysine & Citrulline
Abstract from Patrick Theut's interview with Ivor Cummins plus [Editorial comments]
• Heart disease reversal/prevention requires high serum levels of vitamin K1 and vitamin MK-7 .
• Vitamin K: There is one K1 and 13 [misspoken] K2's. [Edit: The K2's are MK-4 thru MK-13, where the number indicates the length of a molecule chain. So there are 10 of them.]
• K1 is made by plants. It's high in broccoli & Kale.
• K2's come from fermented food products or fermentation by gut bacteria. Carbs feed the wrong type of gut bacteria. Carbs can kill K2 production in your gut. A low carb ketogenic diet is essential for the gut to make K2s. [Edit: Its best to supplement K2's instead of worrying about gut bacteria making them.]
Need for K1, vit-C, and "large fluffy" LDL
• Macrophages inside your cell wall turn into foam cells that cause constant inflammation and the build up of plaque.
• Initially, monocytes patrolling the inside of arteries pull damaged (gluconated or oxidized) lipids and bacteria out of the blood, then become macrophages inside the cell wall.
• Once macrophages job is done they need to be killed, else they become "foam cells", bloated with droplets of cholesterol.
• LDL carries K1 & vit-C. If it has enough K1 and Vit-C it will kill the macrophages that eat it. If there is not enough of either K1 or Vit-C, the macrophages will gorge on LDL and become foam cells.
• This explains why a vegetarian diet high enough in K1 and vit-C could prevent athersclerosis.
• Small LDL cannot carry enough K1, so small LDL will always feed foam cells. You need large fluffy LDL and enough K1 & Vit-C in order to stay healthy.
Need for vit-D & free-T3
• You've got macrophages and foam cells growing and you've got to kill them.
• If you have enough vit-D and enough free T3 (thyroid hormone), there will be proteins on the surface of macrophages that start spinning. This attracts HDL.
Need for MK-7, CoQ10, Micro-RNA, and "large fluffy" HDL
• MK-7 on the HDL locks onto the spinning protein on macrophages or foam cells and locks it down.
• The COQ10 on HDL then binds tightly to the macrophage. Then Micro-RNA from the HDL enters the macrophage (or foam cell) and kills it.
• The HDL then sucks out the fat & cholesterol and when its all bloated it returns to the liver.
• Small HDL cannot carry enough MK7, CoQ10, & Micro-RNA, so it cannot kill foam cells.
• The difference in surface area between small & large HDL is 900%.
• In addition, small HDL cannot orient MK7 & CoQ10 properly to lock onto a macrophage.
Need for magnesium
• A little-known carrier protein is attracted into the artery wall by spinning proteins on macrophages.
• If there is enough magnesium to loosen calcium from muscle cells in the arterial wall, this carrier protein can grab onto the calcium and remove it.
Diet
• Inadequate diet will make LDL & HDL too small
• Will not provide enough K1, vit-C, vit-D, free-T3, MK7, CoQ10, and magnesium.
• High dietary levels create more K1 & vit-C on in LDL
• High dietary levels create a better coating of MK-7, CoQ-10, and Micro-RNA on HDL
• High dietary levels of vit-D and free-T3 create the spinning molecules on macrophages
HDL & LDL will kill & clean out macrophage foam cells with high serum levels of the following:
• LDL
• HDL
• Free T3 between 3.3 & 5.0 pg/ml
• K1, MK-4, & MK-7 koncentrated k supplies all these
• CoQ10
• Vit D3 vit-D, 25-OH between 60 &70
• Vit E
• Vit C Patrick takes 6 g/d, but realizes that is too high for most people
• Magnesium
• Selenium optimum is 200 ug/day (total including food)
• Omega 3's with fat [I take DHA & EPA extracts and avoid fish oil]
• Do not take Calcium or Arginine
• Take Lysine & Citrulline
• One thing to add, since not many would take the time to watch long videos, is that Patrick since almost 2 decades has sieved anything he could find in the available scientific literature (his product page is a testimony to that in respect to K-vitamins only), and is propably the only individual who lab-tested himself the most.
•
• For example, by repeatedly testing undercarboxylated osteocalcin as a proxi for vitamin K2, he found that no high doses of K2 vitamins can be too much, to bring it down to healthier levels. So he know's his own metabolism very well. And many of his recommentations originate from that.
•
• Doing a lot of lab-testing myself (not even nearly as much), I want to point out that there are immense differences in bio-chemical individuality. For example my selenium only came in the optimal range with 370 mcg/d of selenium (incl. food).
• He thinks a carrot a day gives enough preforemed vitamin A. And the liver know's by itself how much is actually needed. In his later video he admits that there are of course genetic differences. I also found carrots not enough, but the need for high doses of preformed vitamin A to raise serum levels to optimal.
• My biome showed I've complete lack in vitamin K1 to K2 metabolizing bacteria, despite being low carb since a decade.
• Only Mg-sulfate IVs, beside supplementing up to 2.5 g of oral elemental Mg a day, finally started to improve my severe Mg-deficiency.
• My rhinitis eats up so much vitamin C for its antihistamine properties, I actually supplemented 24 g/d, and would still do better with more of that stuff.
• So take what it takes, acertained with frequent lab-testing.
•
• Despite obviously being deficient in freeT3 and Mg, I too could halt the mercyless growth in calcification. We are all deferent.
started taking C60oo in early August, 2012, and it has reduced the depth of the wrinkles on my face, as seen in the photos on my profile.
I also noticed fading of two deep scars after Turnbuckle noted that he saw scars fading, likely due to C60oo's potent antioxidant effect improving the health of skin's elastin and collagen.
I assume that the external anti-aging improvements that I've experienced are accompanied by internal improvements.
However, while C60oo appears to reduce the potential for Alzheimer's, cancers, liver inflammation and other potential problems that would limit lifespan, it does not appear to affect cardiovascular issues - blood lipids, etc...
So, with finding out that I have some plaque, I began researching the potential to reverse arterial plaque and improve overall cardiovascular function with a goal of living far past 100 years of age in good health.
I then found a study that showed that high-dose vitamin K2 (MK4) reversed arterial plaque in rats ~37% in just six weeks.
Please see: http://bloodjournal....109/7/2823.long
So, I began taking 5 mg three times a day. I'm raising it to 15 mg, tid, because it is not toxic and I've seen multiple recommendations for 45 mg/day.
I'm also taking vitamin K2 (MK7) 400 mcg/day. It's half life is three days, so I only need to take it once a day.
K1 has beneficial effects for bone, but not plaque, but I am taking 200 mcg/day of it, also.
Turnbuckle enlightened me about xanthohumol's beneficial effects on arterial plaque and HDL, so I'm adding that to my supplement regimen.
Please see: http://www.ncbi.nlm....les/PMC3500296/
I invite you to join in on this pursuit of a healthy, plaque-free cardiovascular system and welcome any insights that you might have.
• like x 4
IP6 (Inositol Hexaphosphate) to the rescue: http://ibdigital.uib...201.dir/201.pdf
"Phytate Inhibits Cardiovascular Calcifications"
---------------------------------------
You might also "Get Thee to a Blood Bank": http://www.healtheir...science-library
Iron status is associated with carotid atherosclerotic plaques in middle-aged adults
Body iron stores and presence of carotid atherosclerosis. Results from the Bruneck Study
Serum ferritin-a novel risk factor in acute myocardial infarction
Ferritin is independently associated with the presence of coronary artery calcium in 12,033 men
I like your K1&K2 dosages, keep it up. But make sure to consume sufficient fat when you take them.
Another great supplement for your purpose is pomegranate extract. LEF sells a very strong one.
Make sure you take sufficient magnesium as D3, K2, Vit A, Magnesium all work in synergy to keep calcium where it needs to be.
Will link this in my part two. Fucking great 👌🏻