Any extracts used in the following article are for non commercial research and educational purposes only and may be subject to copyright from their respective owners.
“Capillary rarefaction is a hallmark of fibrotic disease and reduces blood perfusion and oxygen delivery”1
“Capillary rarefaction is characterized by a reduced density of microvascular networks (arterioles and capillaries) within a vascular bed and is a hallmark of essential hypertension.”2
Published online 11th August ‘22 by Osiaeviet al.3
Abstract
Background
Recent studies have highlighted Coronavirus disease 2019 (COVID-19) as a multisystemic vascular disease. Up to 60% of the patients suffer from long-term sequelae and persistent symptoms even 6 months after the initial infection.
Methods
This prospective, observational study included 58 participants, 27 of whom were long COVID patients with persistent symptoms > 12 weeks after recovery from PCR-confirmed SARS-CoV-2 infection. Fifteen healthy volunteers and a historical cohort of critically ill COVID-19 patients (n = 16) served as controls. All participants underwent sublingual videomicroscopy using sidestream dark field imaging. A newly developed version of Glycocheck™ software was used to quantify vascular density, perfused boundary region (PBR-an inverse variable of endothelial glycocalyx dimensions), red blood cell velocity (VRBC) and the microvascular health score (MVHS™) in sublingual microvessels with diameters 4–25 µm.
Measurements and main results
Although dimensions of the glycocalyx were comparable to those of healthy controls, a µm-precise analysis showed a significant decrease of vascular density, that exclusively affected very small capillaries (D5: − 45.16%; D6: − 35.60%; D7: − 22.79%). Plotting VRBC of capillaries and feed vessels showed that the number of capillaries perfused in long COVID patients was comparable to that of critically ill COVID-19 patients and did not respond adequately to local variations of tissue metabolic demand. MVHS was markedly reduced in the long COVID cohort (healthy 3.87 vs. long COVID 2.72 points; p = 0.002).
Conclusions
Our current data strongly suggest that COVID-19 leaves a persistent capillary rarefication even 18 months after infection. Whether, to what extent, and when the observed damage might be reversible remains unclear.
Study population and study design
This prospective, observational study was conducted from January to December 2021 in the Division of Pulmonary Medicine at University Hospital Münster. We enrolled 27 adults with long-haul COVID (alpha variant) referred to our outpatient clinic due to persistent symptoms > 12 weeks after recovery from PCR-confirmed SARS-CoV-2 infection. Alternative diagnoses were ruled out in all patients by thorough examination. Fifteen apparently healthy, age- and sex-matched volunteers, as well as a historical cohort of hospitalized COVID-19 individuals (n = 16, alpha variant) with critical disease served as negative and positive controls, respectively.
All participants underwent sublingual video microscopy as part of their clinical evaluation by an experienced physician. Exclusion criterion was inflammation of the oral mucosa, which could affect the sublingual microvasculature.
Unfortunately, vaccination status or subclasses were not referenced at all by the authors. This needs to be considered for context, it's important as cases spiked both during and after the rollout, which has implications for LC mediated vascular disease.
What we do know is that by 26th November 2021, in Munster’s federal state of North Rhine-Westphalia, 75.0% of the population had received their first vaccine dose, 71.6% were considered fully vaccinated and 10.3% had received their 3rd booster:
Despite this, by October 2021 German hospitals were seeing a sharp increase in COVID-19 patients.4 Although this was attributed to the unvaccinated, anyone admitted within 14 days of receiving an injection or not having 2 doses were also classed as "unvaccinated" or "not fully vaccinated" at the time.5 The third surge in cases from mid July just should not have happened if the theory of vaccine induced herd immunity was correct. The cure? More of the same of course:
German hospitals see sharp increase in Covid-19 patients
Covid-19 infections have been rising dramatically in Germany - and there are now fears about the increase in people being admitted to intensive care units.
On Thursday Germany reported 28.037 Covid-19 infections in the 24 hours – nearly 12,000 more than a week ago when 16,077 new cases were logged. During the same time period there were 126 Covid-related deaths – the highest number of deaths since June.
The 7-day incidence jumped to 130.2 Covid cases per 100,000 people on Thursday from 118 the previous day. A week ago the incidence was 85.6.
Now health experts are sounding the alarm about the situation in hospitals – and are urging people to get vaccinated.
Chairman of the board of the German Hospital Association, Gerald Gaß, said “We are in a critical pandemic situation.”
Gaß said that the number of patients hospitalised with a Covid infection had risen significantly within a week.
The German Intensive Care register (DIVI) showed that 1,768 Covid-19 patients were in ICU across Germany on October 27th, with 918 receiving ventilation treatment. Doctors say the majority of patients are unvaccinated. There are almost 4,300 Covid patients in regular hospital wards across Germany, reported DPA.
The Robert Koch Institute (RKI) recorded the 7-day incidence of hospitalised cases as 3.07 per 100,000 population on Wednesday – up from 2.13 last Tuesday.
“If this trend continues, we will have 3,000 patients in intensive care again in just two weeks,” Gaß said.
“Even if the hospitals can manage this, it will then not be possible without restricting regular operations,” said the head of the association. It means that doctors in hospitals may have to postpone scheduled, less urgent treatments – like in the previous Covid waves.
At the peak of the pandemic in January 2021, more than 5,700 Corona patients were receiving intensive care treatment.
The Statista graph below shows the number of Covid-19 patients in intensive care in Germany throughout the crisis up to October 26th.
The president of the World Medical Association, Frank Ulrich Montgomery, told the Augsburger Allgemeine on Thursday that those eligible for vaccination who do not get their jabs now are risking their lives, and those of others. “We must try everything in our power to increase vaccination rates,” he said.
“With high vaccination coverage of the population, there are many more mild courses (of Covid-19) – they don’t need to go to hospital, but many unvaccinated people still fall seriously ill.”
Experts have also flagged up staff shortages, in part because of doctors and nurses leaving intensive care medicine due to being overburdened in the pandemic.
Regarding the focus of the paper on long COVID, another study found that vaccination (actually transfection, a gene therapy treatment) only reduces your risk of LC by about 15% after infection.6 This is unfortunate as your likelihood of being infected if transfected more than cancels out this marginal benefit. And that's excluding any post vaccination adverse events not classified as long COVID.
This was from Nov-Dec '21:
“New data from the U.K.’s Office for National Statistics (ONS), says that the triple-vaccinated are 4.5 times more likely to test positive for Omicron than the unvaccinated and that double-vaccinated individuals, were 2.3 times more likely to have Omicron over other variants”.7
Quite a high powered analysis, revealing previous studies were flawed in favour of overstating risks to the unvaccinated:
Vaccination against SARS-CoV-2 lowers the risk of long COVID after infection by only about 15%, according to a study of more than 13 million people. That’s the largest cohort that has yet been used to examine how much vaccines protect against the condition, but it is unlikely to end the uncertainty.
Long COVID — illness that persists for weeks or months after infection with SARS-CoV-2 — has proved difficult to study, not least because the array of symptoms makes it hard to define. Even finding out how common it is has been challenging. Some studies have suggested that it occurs in as many as 30% of people infected with the virus. Buta November study of about 4.5 million people treated at US Department of Veterans Affairs (VA) hospitals suggests that the number is 7% overall and lower than that for those who were not hospitalized.
Another mystery has been whether long COVID is less likely to occur after a breakthrough infection — one in a person who has been vaccinated. In a 25 May study1 in Nature Medicine, nephrologist Ziyad Al-Aly at VA Saint Louis Health Care System in St Louis, Missouri, and his colleagues — the same team that authored the November study — looked at VA health records from January to December 2021, including those of about 34,000 vaccinated people who had breakthrough SARS-CoV-2 infections, 113,000 people who had been infected but not vaccinated and more than 13 million people who had not been infected.
Chinks in the armour
The researchers found that vaccination seemed to reduce the likelihood of long COVID in people who had been infected by only about 15%. That’s in contrast to previous, smaller studies, which have found much higher protection rates. It’s also a departure from another large stud(y), which analysed self-reported data from 1.2 million UK smartphone users and found that two doses of a COVID-19 vaccine halved the risk of long COVID.
The authors of the latest study also compared symptoms such as brain fog and fatigue in vaccinated and unvaccinated people for up to six months after they tested positive for SARS-CoV-2. The team found no difference in type or severity of symptoms between those who had been vaccinated and those who had not. “Those same fingerprints we see in people who have breakthrough infections,” Al-Aly says.
There have been more than 83 million COVID-19 infections in the United States alone, he notes. If even a small percentage of those turn into long COVID, “that’s a staggeringly high number of people affected by a disease that remains mysterious”.
Returning to the research into persistent capillary rarefication in long COVID, the following in vivo assessment of the sublingual microcirculation and glycocalyx dimensions assessments were conducted:
A sidestream dark field (SDF) camera (CapiScope HVCS, KK Technology, Honiton, UK) coupled to the GlycoCheck™ software (Microvascular Health Solutions Inc., Alpine, UT, USA) was used to visualize the sublingual microvasculature and record the movement of red blood cells (RBC) in the microvessels with a diameter (D) of 4–25 µm, as previously described in detail.
A nod to the band Genesis, the glycocalyx has squonk like properties:10
“Making things more complex, the glycocalyx is difficult to stain and visualize, and vanishes when tissue samples are removed for study. In times of shock, the glycocalyx can actually dissipate, making it even harder to find.”
Appearance and behavior
Unlike many mythological creatures, the supposed physical characteristics of the squonk remain unchanged from the original written account, which states:
The squonk is of a very retiring disposition, generally traveling about at twilight and dusk. Because of its misfitting skin, which is covered with warts and moles, it is always unhappy...Hunters who are good at tracking are able to follow a squonk by its tear-stained trail, for the animal weeps constantly. When cornered and escape seems impossible, or when surprised and frightened, it may even dissolve itself in tears.
— William T. Cox, "The Squonk", Fearsome Creatures of the Lumberwoods (1910)
This new research is significant due to the implied risk of public health declining further in the next few months and years, unless significant interventions are conducted now and public health policy is adjusted in favour of safe and effective antiviral therapeutics:
What is the Glycocalyx?
The glycocalyx is a dense layer of sugar molecules and the proteins and fats to which they attach that covers the outer membranes of all the cells in our bodies. The word glycocalyx literally means “sugar coat” (glykys = sweet, kalyx = husk), referring to its carbohydrate composition.
Under an electron microscope, the glycocalyx appears as a thin, fuzzy coating.
While some functions of the glycocalyx have been none for decades, many of its important functions were only been discovered recently.
But the glycocalyx is responsible for many other critical functions:
Regulating circulation to either increase or decrease bloodflow.
Protection: Cushions the plasma membrane of cells protecting against chemical injury.
Regulates Inflammation: Glycocalyx coating on endothelial walls in blood vessels prevents leukocytes from binding.
Fertility: Enables sperm to recognize and bind to eggs.
Embryo development: Guides embryonic cells to destinations in the body.
Immunity against infection: Enables the immune system to recognize and attack. foreign organisms.
Defends against cancer: Changes to the glycocalyx of cancerous cells allow the immune system to recognize and destroy them.
Transplant compatibility: Forms the basis for compatibility of tissue grafts, organ transplants, and blood transfusions.
Cell adhesion: Binds cells together, keeping tissues from falling apart.
Why am I just now hearing about this?
The concept of the glycocalyx dates back to the 1940s, though its structure was impossible to observe at that time. It wasn’t until decades later that electron microscopes became powerful enough to capture the glycocalyx in action.
Making things more complex, the glycocalyx is difficult to stain and visualize, and vanishes when tissue samples are removed for study. In times of shock, the glycocalyx can actually dissipate, making it even harder to find.
The next several decades saw researchers striving to learn more about the glycocalyx and its functions. Some initially theorized that glycocalyx formed a matrix with sieving properties on capillary walls.
Since then, electron microscopy has captured the presence of glycocalyx covering the luminal side (inner side) of capillary walls, though its full function is still unknown.
The glycocalyx is composed primarily of carbohydrates, but that’s not all it contains. It also harbors signaling molecules like cytokines, chemokines, receptors, and growth factors, as well as various enzymes, including superoxide dismutase (SOD). All of these interactions are important for cell function, maintenance, and communication.
So now we know a little about what the glycocalyx is made of. But what does it do?
3 major ways your glycocalyx keeps you healthy
Think of the glycocalyx as a cellular gatekeeper. Just as a gatekeeper keeps intruders out, the glycocalyx guards and protects some of the most delicate cells in your body.
How? Partly by working with the immune system to protect capillaries from invaders. Damaged or toxic cells have often had their glycocalyx stripped away — a testament to its protective abilities.
The benefits of the glycocalyx can be divided into three major categories:
Cardiovascular protection
Hormonal regulation
Increased blood flow
Cardiovascular protection
The glycocalyx protects the capillaries that nourish every cell in our bodies and make up 99% of the circulatory system.
Capillaries are tiny blood vessels that deliver nutrients and remove waste from hard-to-reach places. Capillaries aren’t just hollow pipes that allow blood to flow through them — they’re living, nutrient-exchanging structures. The average adult’s body contains nearly 100,000 miles of them!
With every heartbeat, capillaries transport vital nutrients and oxygen to each cell — and the glycocalyx’s presence facilitates it all.
Here are several more ways this structure protects cardiovascular function:
Its anti-adhesive properties prevent platelets from coagulating and leukocytes from sticking to vascular walls. Prevention of leukocyte adhesion is especially important because leukocytes move around from place to place as needed. The glycocalyx basically adds another layer to this very important immune function.
It’s involved in extracellular signaling (i.e. cell-to-cell communication), which may impair cancer cell growth throughout the cardiovascular system.
Hormonal regulation
The glycocalyx is a significant mediator of nitric oxide (NO) production from endothelial cells.
The glycocalyx seems to subject endothelial cells to just enough pressure to trigger NO production. NO, in turn, “‘informs’ the muscle cells around a blood vessel to contract or relax thus constricting or widening the vessel, so as to regulate the flow of blood.”
Nitric oxide is also involved in regulating the flow of nutrients and oxygen. Without a healthy glycocalyx, this regulation is compromised.
The glycocalyx’s effect on nitric oxide means it has a downstream effect on other hormones, including carbon dioxide, which works with oxygen to optimize your body’s oxygenation levels.
Increased blood flow
The glycocalyx can also increase blood flow, partially thanks to its ability to mediate nitric oxide. There are other pro-circulation benefits, too:
Allows the body to engage more of its capillaries, delivering more nutrients to and/or removing waste from organs as needed.
Protects the endothelial cells from direct pressure caused by blood flow, helping to regulate their shape.
Regulates the permeability of blood vessels, preventing an increased influx of proteins, water, and other blood-borne elements like low-density lipoprotein (“bad” cholesterol).
Why the glycocalyx health is so important
Simply put, the glycocalyx is essential for protecting the health of the circulatory system.
Circulatory health, in turn, has many downstream benefits.
There’s a potential problem, though: the glycocalyx is delicate. And while it degrades naturally over time or during certain stressful events, this process can be exacerbated by other factors, including:
High-carbohydrate diet
Lack of exercise
Genetic factors
Chronic inflammation
High blood pressure
Smoking
Trauma
ARDS
COVID-19
Damage to the glycocalyx can also cause shedding of the antioxidant enzyme SOD, which converts reactive oxygen species (ROS) to hydrogen peroxide. Without SOD, the balance of the endothelium is shifted to a pro-oxidant, hyper-inflammatory state.
Furthermore, damage to the glycocalyx can lead to further trauma to the vascular endothelium, leading to leaky capillaries that may lose function and/or die off.
What diseases have been linked to glycocalyx damage?
Without the vital nutrients and oxygen supplied by capillaries, tissues and vital organs starve. Impaired tissue perfusion has been repeatedly shown to be the root cause of major diseases and health complications, including but not limited to:
Sepsis
Diabetes (type 1 and type 2)
Heart failure
Lung disease
Kidney disease
Stroke
Dementia
Early-onset preeclampsia
Inflammatory disorders
Cancer progression
Unfortunately, medical researchers estimate that a large percentage of the world’s population is affected by glycocalyx breakdown.
Atherosclerosis
As we discussed earlier, inflammation can trigger the endothelial glycocalyx to dissipate, which only promotes further inflammation.
This dissipation can allow ‘bad’ cholesterol to enter the bloodstream, become oxidized, and form a lipid layer. This further promotes the shedding of the glycocalyx. Eventually, leukocytes and other types of white blood cells called monocytes and lymphocytes begin to infiltrate the intima or innermost coating of the endothelium.
Once monocytes enter the intima, they become macrophages, which consume oxidized LDL cholesterol to form a fibrous cap, thereby stabilizing the plaque. If this fibrous cap is disturbed, it can lead to cardiovascular events like heart attacks and strokes.
The steps described above are highly simplified versions of the complex process that is atherosclerosis. However, it’s clear that the glycocalyx plays a crucial role in resisting the build-up of plaques and keeping blood vessels healthy.
Why a high-carb diet damages the glycocalyx
Eating a diet high in carbohydrates can spell trouble by causing spikes in blood sugar levels.
In one research study, only 6 hours of exposure to high blood sugar reduced glycocalyx volume by a whopping 50%. The research team credited high blood sugar’s damaging effects to increased oxidation and inflammation. This was confirmed when they infused glucose with a strong antioxidant and the glycocalyx volume remained unchanged.
Considering that glycocalyx damage is thought to be the “first step” towards atherosclerotic problems of all kinds, the speed with which high glucose can reduce glycocalyx volume is eye-opening.
High blood sugar may also cause the mitochondria (the energy factories in each cell) to overproduce too many free radicals, including reactive oxygen species (ROS). ROS are key signaling molecules that regulate the natural inflammatory responses in our bodies.
In a healthy system, antioxidants quench ROS when they are no longer necessary, but too many and chronically high ROS can lead to oxidative stress, DNA damage, and eventually chronic diseases like cancer, and heart disease, among many others.
In short, a high-carb diet can devastate the glycocalyx — and your health.
How to eat for your glycocalyx
Given its significance, the glycocalyx may serve as a promising target in the treatment of chronic diseases. Researchers have been looking into ways to regenerate and/or protect the glycocalyx.
In the absence of any new breakthroughs, however, we know of one surefire way to protect your glycocalyx from breakdown: eat a low-carb, high-fat diet, ideally a carnivore diet.
When carbohydrate intake is reduced, blood sugar levels will stabilize, preventing damage to the glycocalyx and its sensitive glycolipids and glycoproteins.
Certain supplements may also help maintain a healthy glycocalyx. These include:
Hyaluronic acid
N-acetyl cysteine (NAC)
Chondroitin sulfate
Nitric oxide boosters
Activated protein C
Hydrocortisone
Progesterone
While these supplements may work, they shouldn’t be used as a bandaid solution. Consider taking the holistic route — eating a nourishing, anti-inflammatory diet — first. And always consult a healthcare professional before making drastic changes.
The Takeaway
Scientists are continuing to learn just how important the glycocalyx is for the cardiovascular system and for general health.
The more they learn, the clearer the dangers of regular sugar and starch consumption become. As we’ve seen, hyperglycemia can prematurely degrade the glycocalyx covering of capillaries. This simple-yet-serious occurrence may very well explain the high rates of cardiovascular diseases seen in our country.
Thankfully, protecting yourself is as easy as protecting your glycocalyx. You can get started by reducing your consumption of processed carbohydrates and focusing on healthy fats and proteins. Stress-reducing practices like walking, yoga, mindful eating, and meditation may also help.
Take care of your glycocalyx, and it’ll take care of you.
In the study of review, the following parameters were calculated:
Perfused boundary region (PBR, in µm), an estimate of the dynamic lateral movement of RBCs into the permeable part of the endothelial glycocalyx layer. The higher the PBR values, the more diminished the glycocalyx dimensions.
RBC velocity (VRBC, in µm/s), calculated by dividing longitudinal RBC displacement by the time between video frames.
Capillary density (in 10−2 mm/mm2), defined as the vascular density of vessels with a diameter ≤ the diameter of a single RBC (D ~ 7–8 µm [9]); capillary density D < 7 µm.
Microvascular health score (MVHS™, in points), a novel score that combines microcirculatory impairment and eGC dimensions; higher values indicate healthier microvasculature. The MVHS is calculated by dividing capillary blood volume (CBV; i.e., capillary density multiplied by segment-specific capillary cross-sectional area (ππ * radius2) by the PBR value.
Capillary recruitment (CR, in %), an estimate of the microvascular ability to recruit additional capillaries under stress conditions. CR can be derived per group by the slope of the relationship between VRBC in the capillaries (≤ 7 µm) and in the feed vessels (≥ 10 µm) and is defined as 1 − slope [VRBC (D ≤ 7 µm), VRBC (D ≥ 10 µm)].
The number of participants was comparatively small at 27, but age and comorbidity matching was good, to help minimise confounding factors. Either way the reduction in capillary density in LC patients was a highly significant 41%:
Results
The long COVID cohort encompasses 27 adult patients, studied after a median (IQR) duration of 541 (305–569) days post-COVID-19 infection. The patients (55.5% men), aged 53 (39–51) years and had a median Charlson Comorbidity Index (CCI) of 0 (Table 1).
Long COVID patients showed a significantly lower vascular density compared to healthy controls. Specifically, a µm-precise analysis showed the decrease in vascular density to exclusively affect very small capillaries (D5: − 45.16%; D6: − 35.60%; D7: − 22.79%) (Fig. 1A). Compared with healthy controls, the capillary density of long COVID patients was decreased by 41% (Fig. 1B). However, glycocalyx dimensions did not differ from those of the healthy volunteers (Fig. 1C), even in a µm-precise analysis (data not shown). Upon recombination of these indices into MVHS™, severe impairment of the microcirculation in individuals with long COVID [healthy 3.87 (3.15–4.87) vs. long COVID 2.72 (2.21–3.01) points, p = 0.002; Fig. 1D] is emphasized. In a pooled analysis of the measurements of all study participants (n = 58), disease group (healthy controls, long COVID, historical acute COVID-19 cohort) was the only variable associated with the MVHS™. This association remained significant after adjustment for age, sex, CCI and BMI (β = − 0.52, p < 0.001).
Microvascular phenotyping by quantitative sublingual video microscopy. A Median and IQR values of vascular density of healthy controls (green), and long-haul COVID-19 (orange) patients based on the diameter class from 4 to 25 µm. A historical cohort of hospitalized COVID-19 patients (pink) is shown for reference. *q < 0.05, **q < 0.01, ***q < 0.001. B–D Boxplots of B capillary density (D4–6 µm) and C PBR4–25 µm and D MVHS™ of healthy controls (green), long-haul COVID individuals (orange), COVID-19 patients (pink). COVID-19 Coronavirus disease 2019, CR capillary recruitment, D diameter, PBR perfused boundary region, RBC red blood cell, VRBC red blood cell velocity, MVHS™ microvascular health score
They measured red blood cell velocity (VRBC) and found that it increased significantly in LC patients, which the authors suggest is a compensatory measure. But further increases in tissue metabolic demand did not lead to an increase in the number of perfused capillaries, unlike the healthy control group. What this means is that you may quickly hit a wall of muscle cramps or exhaustion during heavy exercise, and it can have an impact on neurocognitive function (aka brain fog).
CBV: Capillary blood volume.
CR: Capillary recruitment.
We previously used a ratio derived from VRBC in feed vessels relative to capillaries (VRBC (D ≥ 10 µm)/VRBC (D ≤ 7 µm) to calculate capillary recruitment (see above) [8]. Analysis of RBC velocity (VRBC) revealed significant differences between all three study groups, both in VRBC4–7 µm (Fig. 2A) and in VRBC≥10 µm (Fig. 2B). Surprisingly, VRBC was significantly higher in capillaries as well as feed vessels in long COVID patients compared with healthy individuals (VRBC4–7 µm: 114.9 [102.7–134.4] vs. 103 [89.87–112.6] µm/s, p = 0.01; VRBC≥10 µm: 124 [119.6–132.2] vs. 100.6 [93.95–110.1] µm/s; p < 0.0001). Plotting VRBC≥10 µm vs. VRBC4–7 µm revealed a strong dependency between capillaries and feed vessels in both, individuals with long COVID and hospitalized COVID-19 patients (long COVID: R2 = 0.29, p = 0.004; COVID-19: R2 = 0.45, p < 0.0001), indicating impaired capillary (de-)recruitment in these groups. In contrast, capillary VRBC was constant in healthy controls, indicating functioning (de-)recruitment of CBV associated with changes of feed vessel blood flow in healthy subjects (R2 = 0.12; p = 0.23) (Fig. 2C). CR was 29% in long COVID, 17% in COVID-19 patients and 70% in healthy volunteers (Fig. 2D). This finding suggests that the number of perfused capillaries in long COVID remains fixed and does not adequately respond to local variations of tissue metabolic demand. Associating the different symptoms with the capillary density in an explorative manner, revealed a statistically significant capillary rarefication in long COVID individuals presenting with neurocognitive dysfunction (38.50 [25.93–44.41] vs. 49.77 [43.40–64.30], p = 0.049; Supp. Table 1).
Red blood cell velocities and recruitment capacity. A, B Boxplots of A VRBC in capillaries (D ≤ 7 µm) and B VRBC in feed vessels (D ≥ 10 µm) of long COVID individuals (orange), hospitalized COVID-19 patients (pink) and healthy controls (green). C Scatter dot plots and simple linear regression (slope) with 95% confidence intervals of VRBC in capillaries plotted against VRBC in feed vessels of long COVID individuals (orange), hospitalized COVID-19 patients (pink) and healthy controls (green). D Bar charts showing the capillary recruitment (CR), defined as 1 − slope [VRBC (D ≤ 7 µm) vs. VRBC (D ≥ 10 µm)] per group
MVHS: Microvascular health score.
Perhaps most alarming was that the MVHS declined equally regardless of clinical outcome at the time, and the effects appear to be irreversible:
By further analyses, long COVID individuals showed no significant difference in MVHS or capillary density irrespective of outpatient or inpatient care—regardless of need for oxygen supplementation, during acute COVID-19 infection (Fig. 3A, C; Table 2). To assess recovery of microcirculatory parameters over time, we plotted MVHS and capillary density against the length of time between initial infection and presentation to the long COVID study outpatient clinic. Interestingly, no improvement of these parameters was documented, assuming capillary loss due to COVID-19 might be irreversible (Fig. 3B, D).
Sensitivity analyses in individuals with long COVID. A, C Boxplot of (A) MVHS and (C) density of long COVID individuals based on the clinical course of their acute COVID-19 infection. B, D Scatter dot plots and simple linear regression (slope) with 95% confidence intervals of (B) MVHS and (D) capillary density plotted against the days after the COVID-19 infection. COVID-19 Coronavirus disease 2019, MVHS microvascular health score, ns not significant
By “persistent immunological abnormalities” they mean “induced autoimmunity and putative viral reservoirs”.11
As per my literature review, spike protein from transfection puts you at a much greater relative risk of this due to repeated booster dosing and re-infections.12
Consider also that myocardial microcirculation is being compromised here too, which may well be life shortening if sustained without therapeutic intervention to try to mitigate the damage:
To what extent this capillary rarefication is mechanical or/and functional remains unclear. Pretorius et al. revealed common clotting pathologies in plasma of acute and long COVID patients, further supporting the existence of persistent microthrombi. Besides that, an insufficient recovery of the initial inflammation accompanied by persistent immunological abnormalities might also be responsible for the observed capillary impairment [6]. Previous studies focusing on the pulmonary, exertional and myocardial microcirculation of long COVID individuals showed local impairment of the microcirculation. Our data complement these findings and point to a systemic and long-lasting capillary rarefication. This finding could possibly explain the functional impairment observed in long COVID syndrome. A subtle but exciting feature of the long-haul COVID group is, that the RBC velocities are the highest in our long COVID cohort (indicated by parallel shift of the slope to the upper right). We speculate that this finding could represent a compensatory mechanism to meet metabolic demands. Considering that the measurements were taken at physical rest, it is quite conceivable that this presumed compensatory mechanism is exhausted more quickly during exertion than in healthy individuals, possibly explaining long COVID symptoms. Here, we previously were able to present data on persistent exertional impairment caused by (micro-) circulatory reduction of the oxygen pulse during sequential, bicycle cardiopulmonary exercise test.
In the TUN-EndCOV Study, Charfeddine et al. reported a significant microvascular and endothelial dysfunction in a post-COVID cohort, evaluated by finger thermal monitoring after occlusion and reperfusion of the hand. Specifically, long COVID individuals showed a higher endothelial quality index and a slower response to the reperfusion phase. This finding was partially reversed after oral intake of sulodexide, a highly purified mixture of glycosaminoglycans, that includes fast-moving heparin and dermatan sulfate. Indeed, our data imply that long COVID individuals respond inadequately to local variations of tissue metabolic demand. Therefore, it is intriguing to speculate, that this delayed response after reperfusion might be the functional result of the observed capillary rarefication and the impaired capillary recruitment of the remaining capillaries in the long COVID individuals.
They report on a recovery of glycocalyx dimensions after about 18 months. But if you get transfected or re-infected in that period then any recovery would be reversed, according to known spike protein mediated pathophysiology, which will be considered later.
“Primum non nocere. First, do no harm”.
Regarding endothelial glycocalyx, previous studies have shown a trend towards partial recovery of the glycocalyx in the first months following the acute infection. Ikonomidis et al. evaluated PBR values of post-COVID individuals 12 months after infection, showing persistent glycocalyx impairment. Here we demonstrate completely restored glycocalyx dimensions in long COVID individuals about 1.5 years after the acute infection. Mechanistically, glycocalyx thinning is tightly controlled, among others by the endothelium specific Angiopoietin(Angpt)/Tie2 system. We have shown that Angpt-2 activates heparanase release from the endothelium which leads to enzymatic degradation of the endothelial glycocalyx. Indeed, we and others showed that both heparanase and Angpt-2 levels are elevated during the acute infection. Recent studies in long COVID reported normalized heparanase and Angpt-2 levels, consistent with restored glycocalyx dimensions. In this regard, our data once again highlight differential regulation of microcirculation parameters and glycocalyx dimensions, a finding we first observed in critically ill patients with bacterial sepsis.
We acknowledge some limitations. First, our pilot study had a small sample size and is mostly hypothesis-generating. Our study is observational and does not demonstrate causality between microvascular damage and post-COVID symptoms. Second, we included a historical cohort to better understand the microvascular changes. However, all microvascular parameters were assessed under the same conditions using the same GlycoCheck camera by the same experienced investigator. Third, plasma samples are not available in our long COVID patients, so we cannot perform further analyses of endothelial markers. Fourth, the capillary recruitment was calculated post-hoc in a per group analysis; meanwhile a per individual analysis has become available which might provide even more accurate results.
Conclusion and outlook
Microvascular impairment appears to play a crucial role in the pathophysiology of both COVID-19 and post-COVID sequelae. In summary, our current data strongly suggest that COVID-19 leaves a persistent capillary rarefication even 18 months after infection. Whether, to what extent, and when the observed damage might be reversible remains unclear. If capillary rarefaction were to persist, additional cardiovascular disease (e.g., diabetes, hypertension) will be less well compensated and become symptomatic much earlier. It therefore seems important to understand the mechanisms of capillary rarefication in long COVID in detail. Here, we see a useful application for our µm-wise quantitative video microscopy in characterizing study patients (enrichment) and monitoring response to potential therapies.
No conflict of interest declaration was given.
Pathophysiology - further research
This paper by Leif Østergaard from 2021 discusses the positive feedback loop induced by microvascular damage:13
Abstract
Corona virus disease 2019 (COVID‐19) causes symptoms from multiple organs after infection by severe acute respiratory syndrome corona virus 2 (SARS CoV‐2). They range from early, low blood oxygen levels (hypoxemia) without breathlessness (“silent hypoxia”), delirium, rashes, and loss of smell (anosmia), to persisting chest pain, muscle weakness and ‐pain, fatigue, confusion, memory problems and difficulty to concentrate (“brain fog”), mood changes, and unexpected onset of hypertension or diabetes. SARS CoV‐2 affects the microcirculation, causing endothelial cell swelling and damage (endotheliitis), microscopic blood clots (microthrombosis), capillary congestion, and damage to pericytes that are integral to capillary integrity and barrier function, tissue repair (angiogenesis), and scar formation. Similar to other instances of critical illness, COVID‐19 is also associated with elevated cytokine levels in the systemic circulation. This review examines how capillary damage and inflammation may contribute to these acute and persisting COVID‐19 symptoms by interfering with blood and tissue oxygenation and with brain function. Undetectable by current diagnostic methods, capillary flow disturbances limit oxygen diffusion exchange in lungs and tissue and may therefore cause hypoxemia and tissue hypoxia. The review analyzes the combined effects of COVID‐19‐related capillary damage, pre‐existing microvascular changes, and upstream vascular tone on tissue oxygenation in key organs. It identifies a vicious cycle, as infection‐ and hypoxia‐related inflammation cause capillary function to deteriorate, which in turn accelerates hypoxia‐related inflammation and tissue damage. Finally, the review addresses the effects of low oxygen and high cytokine levels in brain tissue on neurotransmitter synthesis and mood. Methods to assess capillary functions in human organs and therapeutic means to protect capillary functions and stimulate capillary bed repair may prove important for the individualized management of COVID‐19 patients and targeted rehabilitation strategies.
COVID‐19‐related microvascular damage and inflammation may cause tissue hypoxia via transit‐time effects and disturb neurotransmitter synthesis in the brain. The duration of COVID‐19 symptoms and the long‐term health effects of SARS‐CoV‐2 infection may rely on whether disease‐related capillary damage is reversible.
Not just from COVID-19 viral infection, but transfection mediated spike protein also upregulates tumor necrosis factor alpha (TNF‐α) and induces clotting and consequential hypoxic tissue damage via multiple signalling pathways.
4.3. Glycocalyx
The capillary endothelium's luminal surface is covered by a 0.3–0.6 µm thick glycocalyx (Vink & Duling, 1996), a matrix that acts as a fluid barrier (Haaren et al., 2003) and impacts erythrocytes’ passage (Secomb et al., 1998). In critical illness, partly due to elevated TNF‐α levels, the glycocalyx is shed (Henry & Duling, 2000; Nieuwdorp et al., 2009), exposing cellular adhesions molecules that interact with immune cells to facilitate their extravasation. Cortisol protects the endothelial glycocalyx against TNF‐α induced shedding (Chappell et al., 2009). Glycocalyx shedding profoundly affects microvascular resistance and capillary hemodynamics (Cabrales et al., 2007; Lipowsky et al., 2011). In newly intubated COVID‐19 patients, Stahl et al. found evidence of glycocalyx shedding before signs of any EC injury and verified glycocalyx thinning by sublingual optical imaging (Stahl et al., 2020).
4.4. Capillary obstruction by neutrophils (“stalled flow”)
Neutrophils are involved in the immune response to SARS‐CoV‐2 infection. Much larger than erythrocytes and the average capillary diameter, they may occlude capillaries for several seconds, and especially when activated, cause significant capillary flow disturbances (Harris & Skalak, 1993). Neutrophil adhesion in brain capillaries impairs memory function and causes sizeable reductions in cerebral blood flow in animal models (Cruz Hernandez et al., 2019). The adhesion of hyper‐activated neutrophils to capillaries within the lungs, brain, heart, and other organs may therefore contribute to the poor prognosis of some COVID‐19 patients (Wang et al., 2020).
4.5. Microthrombosis
Capillary congestion and microthrombosis in the absence of upstream thrombi are common findings in the lungs of patients who died from COVID‐19‐related respiratory failure (Ackermann et al., 2020; Carsana et al., 2020). Microthrombosis has also been observed in the skin (Colmenero et al., 2020) and the kidney's glomerular capillaries (Hanley et al., 2020).
6. HYPOXIA, INFLAMMATION, AND CAPILLARY FUNCTION
COVID‐19, like other instances of critical illness that involve systemic hypoxia (Kox et al., 2020; Sinha et al., 2020), is associated with cytokine release. TNF‐α and other inflammatory factors are known to damage the glycocalyx and impair endothelial functions (Liu et al., 2020; Zhang et al., 2020). This immune response is likely to contribute to capillary dysfunction and reduced tO2 in affected tissue, where cellular oxygen sensors, in turn, activate hypoxia‐inducible transcription factor (HIF) and the transcription of genes that help tissue adapt to hypoxia (Eltzschig & Carmeliet, 2011). Notably, hypoxia also activates the production of nuclear factor kappa beta (NF‐κB), the master‐switch for the transcription of genes that elicit inflammatory responses (Eltzschig & Carmeliet, 2011). Figure Figure33 illustrates how hypoxia‐induced inflammation, by adding further to capillary dysfunction and hypoxia, may constitute a vicious cycle in severe COVID‐19.
Interactions between capillary function, inflammation, hypoxia, and neurotransmission. The expression of ACE2 and other SARS‐CoV‐2 entry factors on parenchymal cells and observations of infected cells in biopsy material hold important clues to understand COVID‐19‐related organ damage
I reviewed the related hypoxic kynurenine pathway and Warburg effect pathology previously.14
Barhoumi et al (2021) investigated spike protein induced apoptosis, inflammatory and oxidative stress responses:15
Any ACE2 expressing cells, including endothelial linings of capillaries, will be at risk from this signalling pathway:16
Abstract
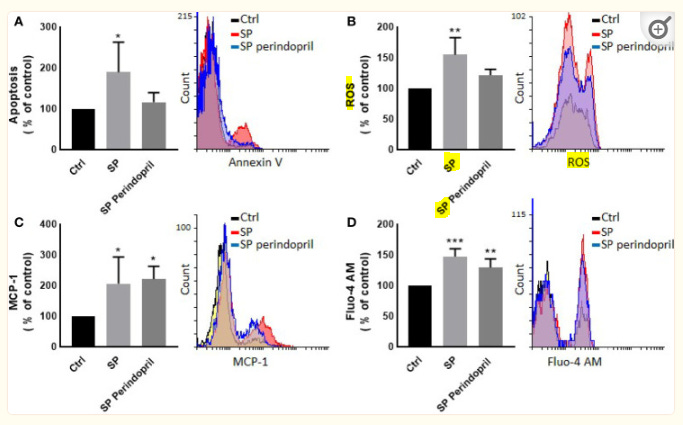
A purified spike (S) glycoprotein of severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) coronavirus was used to study its effects on THP-1 macrophages, peripheral blood mononuclear cells (PBMCs), and HUVEC cells. The S protein mediates the entry of SARS-CoV-2 into cells through binding to the angiotensin-converting enzyme 2 (ACE2) receptors. We measured the viability, intracellular cytokine release, oxidative stress, proinflammatory markers, and THP-1-like macrophage polarization. We observed an increase in apoptosis, ROS generation, MCP-1, and intracellular calcium expression in the THP-1 macrophages. Stimulation with the S protein polarizes the THP-1 macrophages towards proinflammatory futures with an increase in the TNFα and MHC-II M1-like phenotype markers. Treating the cells with an ACE inhibitor, perindopril, at 100 µM reduced apoptosis, ROS, and MHC-II expression induced by S protein. We analyzed the sensitivity of the HUVEC cells after the exposure to a conditioned media (CM) of THP-1 macrophages stimulated with the S protein. The CM induced endothelial cell apoptosis and MCP-1 expression. Treatment with perindopril reduced these effects. However, the direct stimulation of the HUVEC cells with the S protein, slightly increased HIF1α and MCP-1 expression, which was significantly increased by the ACE inhibitor treatment. The S protein stimulation induced ROS generation and changed the mitogenic responses of the PBMCs through the upregulation of TNFα and interleukin (IL)-17 cytokine expression. These effects were reduced by the perindopril (100 µM) treatment. Proteomic analysis of the S protein stimulated THP-1 macrophages with or without perindopril (100 µM) exposed more than 400 differentially regulated proteins. Our results provide a mechanistic analysis suggesting that the blood and vascular components could be activated directly through S protein systemically present in the circulation and that the activation of the local renin angiotensin system may be partially involved in this process.
Graphical: Suggested pathways that might be involved at least in part in S protein inducing activation of inflammatory markers (red narrow) and angiotensin-converting enzyme inhibitor (ACEi) modulation of this process (green narrow).
Note the reactive oxygen species (ROS) surge of oxidative free radicals in response, which are highly pathogenic to cells and mitochondrial DNA in particular, and correlate positively with neurodegenerative disease17, endothelial damage, cardiovascular disease, metabolic disorders18 and carcinogenesis amongst others19.
Effects of spike (S) protein on cell viability and inflammatory markers. Thp-1 macrophages were mock treated (Ctrl), stimulated with S protein (SP) (100 nM) or SP with ACE inhibitor (SP Perindopril, 100 µM), perindopril, added 2 h before SP. (A) Apoptosis was measured by Annexin V binding essay. (B) Oxidative stress was evaluated by reactive oxygen species (ROS) expression. (C) Monocyte chemoattractant protein-1 (MCP-1) expression essay. (D) Fluo4 fluorescence as a function of cytosolic free Ca2+ intracellular calcium was performed using flow cytometry analysis. The data are presented as mean ± SEM, n = 4–5. Ordinary one-way ANOVA followed by Dunnett’s test: *p < 0.05, **p < 0.01,***p < 0.001.
I reviewed the pathological effects of interleukin-17 separately.20
Effects of spike (S) protein stimulation on PBMCs. PBMCs were mock treated controls (Ctrl), stimulated with S protein (SP) (100 µM), or SP with ACE inhibitor (SP Perindopril, 100 µM), perindopril, added 2 h before SP. Flow cytometry analysis was performed for measurement of (A) expression of TNFα in PBMCs, (B) expression of TNFα on CD4 positive T cells, (C) expression of intracellular IL-17 on CD4 T cells, and (D) ROS production on PBMCs. The data are presented as mean ± SEM, n = 4. Ordinary one-way ANOVA followed by Dunnett’s test: *p < 0.05, **p < 0.01
It is known that ROS play crucial roles in the physiological processes in cells. The dysregulation of the redox balance increase free radicals, which cause damage and alters several cellular functions (45, 46). During the ARDS phase of the infection in COVID-19, ROS induced cell damage and altered signaling pathways (45). The exposure of the PBMCs to the S protein for 24 h significantly increased the ROS levels compared to the mock-treated controls. The treatment of perindopril did not change this effect (Figure 3D).
Inflammation without antiviral interferon immune response upregulation:
IFNγ and TNFα are crucial cytokines involved in innate and adaptive immunity, and their dysfunction or abnormal release is a key modulator for macrophage and immune cell activation and viral clearance. To evaluate the potential immune response of lymphocytes, S protein treatment before mitogen stimulation induced the upregulation of the intracellular TNFα and IL-17 in whole PBMCs and CD4 T helper cells. This effect is possibly due to the activation of monocytes that sequentially activate T-cell subpopulations differentially, as we did not find any effect on the CD8 cytotoxic T cells. We also did not find a significant effect on IFNγ expression on the CD4, CD8, or NK cells, suggesting that the S protein binding to ACE2 may not be a stimulator for any pathway related to IFNγ release following mitogen activation.
The M1 response is inflammatory and tumorigenic:21
Collectively, our findings highlighted the evidence that S glycoprotein in vitro stimulation potentially promote the differential activation of THP-1-like macrophages, HUVECs, and PBMCs. S protein induces ROS generation in PBMCs and THP-1-like macrophages, most probably due to the Ero1α protein upregulation. S protein induction polarizes the THP-1-like macrophages toward the M1-like phenotype and modulates intracellular TNFα and IL-17 expression in the activated PBMCs and HIF1α and MCP-1 in HUVEC cells. These effects were blunted partially after treating the cells with an ACE inhibitor. These results indicated that the proinflammatory mediators could be stimulated by S protein interaction with immune and endothelial cells and that the SARS-CoV-2 virus infection may differentially activate host cells independently, which potentially explain the variety of clinical phenotypes and severity progress of COVID-19. In view of the fact that our study was in vitro using spike protein, more investigations using live virus or an animal model are highly valuable for targeting this area that may be of interest for the diagnosis and treatment of COVID-19.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
In 2021, Khan et al published research into how the spike protein, even in the absence of the virus, induces inflammation via TLR2-dependent activation of the NF-κB pathway.22
Note that although glycocalyx damaging TNFa is upregulated, as discussed previously antiviral interferons are not. This is to be expected as viral survival would be impaired by the resultant positive feedback loop:
Abstract
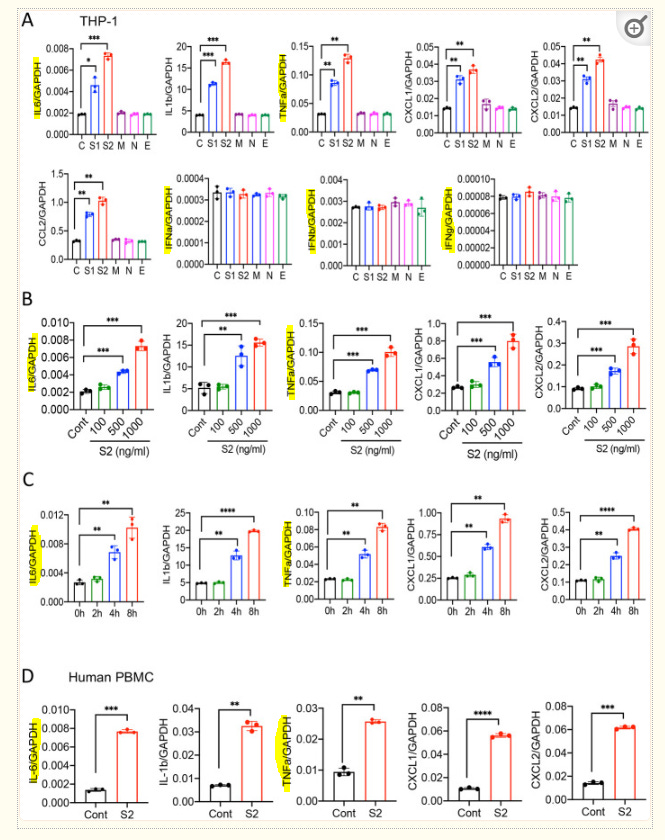
Pathogenesis of COVID-19 is associated with a hyperinflammatory response; however, the precise mechanism of SARS-CoV-2-induced inflammation is poorly understood. Here we investigated direct inflammatory functions of major structural proteins of SARS-CoV-2. We observed that spike (S) protein potently induces inflammatory cytokines and chemokines including IL-6, IL-1β, TNFα, CXCL1, CXCL2, and CCL2, but not IFNs in human and mouse macrophages. No such inflammatory response was observed in response to membrane (M), envelope (E), and neucleocapsid (N) proteins. When stimulated with extracellular S protein, human lung epithelial cells A549 also produce inflammatory cytokines and chemokines. Interestingly, epithelial cells expressing S protein intracellularly are non-inflammatory, but elicit an inflammatory response in macrophages when co-cultured. Biochemical studies revealed that S protein triggers inflammation via activation of the NF-κB pathway in a MyD88-dependent manner. Further, such an activation of the NF-κB pathway is abrogated in Tlr2-deficient macrophages. Consistently, administration of S protein induces IL-6, TNF-α, and IL-1β in wild-type, but not Tlr2-deficient mice. Together these data reveal a mechanism for the cytokine storm during SARS-CoV-2 infection and suggest that TLR2 could be a potential therapeutic target for COVID-19.
Note the upregulation of interleukin-6 (Il-6) and tumor necrosis factor alpha (TNFa), both of which are pro tumorigenic as well as pro-inflammation.23
Interleukin-1 β (IL-1 β) is a major cytokine involved in monocyte activation and activation of proinflammatory signaling pathways in peripheral tissues and brain.24
The CXCL’s are chemokines, small peptides that act as chemoattractants for several immune cells. This is part of the inflammatory response which can cause necrosis of various organ tissue cells, including those of the cardiac pericardium and myocardium.
If this occurs in the absence of a pathogen then it may be considered to be an induced autoimmune disease response.
SARS-CoV-2 S protein induces cytokines and chemokines in macrophages and monocytes.
(A) Human monocytic cells THP1-derived macrophages were stimulated with recombinant S1, S2, M, N, and E proteins of SARS-CoV-2 at a concentration of 500 ng/ml. Four hours post stimulation, the expression of IL6, IL1b, TNFa, CXCL1, CXCL2, CCl2, IFNa, IFNb, and IFNg was measured by real-time qPCR. (B) THP-1 cells were stimulated with S2 protein (500ng/ml). At indicated times, RNA was isolated and measured for IL6, IL1b, TNFa, CXCL1, and CXCL2 by real-time qPCR. (C) THP-1 cells were stimulated with S2 protein at various concentration for 4 h, and measured the indicated cytokines by real-time qPCR. (D) Human peripheral blood mononuclear cells (PBMC) were incubated with S2 protein for 4h. The expression of IL6, IL1b, TNFa, CXCL1, and CXCL2 was measured by real-time qPCR. Data represent mean ± SD (n=3); *p < 0.05, **p < 0.001, ***p < 0.0001, ****p < 0.00001 by unpaired Student’s t test. Experiments were repeated two times and data of a representative experiment is presented.
Similar results were obtained with lung epithelial cells:
Lung epithelial cells produce inflammatory molecules in response to SARS-CoV-2 S protein.
(A) A549 cells were incubated with SARS-CoV-2 S2 (500 ng/ml) protein for 12 and 24h. RNA was isolated and measured for the expression of inflammatory cytokines and chemokines. (B-C) SARS-CoV-2 S protein was overexpressed in HEK293T and A549 cells. 48h following transfection with expression plasmids, the mRNA levels of IL6, IL1b, TNFa, CXCL1, and CXCL2 were measured. Data represent mean ± SD (n=3). Experiments were repeated two times and data of a representative experiment is presented.
S protein activates the NF-κB pathway
Inflammatory genes are transcriptionally regulated by transcription factors that are activated by signaling pathways such as NF-κB, MAPK, STAT3, and AKT. To obtain further insight into how S protein induce the expression of inflammatory mediators, we stimulated THP1 cells with S2 protein. Cell lysates collected at various times following stimulation were analyzed for the activation of these inflammatory pathways by western blotting. As shown in Figure 4A, P65 and IκB were phosphorylated in cells treated with S2 protein. The activation of NF-κB pathway is often accompanied by the activation of MAPK pathways, including ERK, P38 and JNK. Surprisingly, there was no activation of ERK and JNK in S2 stimulated cells (Figure 4A). There was no activation of the AKT pathway as well (Figure 4A), while STAT3 was phosphorylated at 2h following stimulation (Figure 4A). Inflammatory cytokines, such as IL-6, can activate STAT3; thus, the observed activation of STAT3 could be a secondary response of S protein-mediated activation of the NF-κB pathway. S2 protein also activated the NF-κB and STAT3 pathways in A549 cells (Figure 4B). To confirm that S protein-induced inflammation was NF-κB dependent, we inhibited the NF-κB pathway using Sc514, an inhibitor of IKKβ, during stimulation with S protein. As expected, inhibition of the NF-κB pathway abrogated inflammatory responses in S protein-stimulated macrophages (Figure 4C and and D).
(A-B) THP1 and A549 cells were stimulated with S2 (500ng/ml) for indicated time points. Phosphorylation of P65, IκBα, ERK, JNK, STAT3, and AKT was measured by western blotting. (C-D) THP1 cells were stimulated by SARS-CoV-2 S2 protein (500ng/ml) in the presence or absence of IKKβ inhibitor sc514. (B) Phosphorylation of P65 and IκBα was measured by Western blotting. (E) The expression of IL6, IL1b and TNFa in stimulated THP1 cells was measured by real-time qPCR. Data represent mean ± SD (n=3); *p < 0.05, **p < 0.001, ***p < 0.0001 by unpaired Student’s t test. Experiments were repeated two times and data of a representative experiment is presented.
NB Protein phosphorylation is an important cellular regulatory mechanism as many enzymes and receptors are activated/deactivated by phosphorylation and dephosphorylation events, by means of kinases and phosphatases.25
Toll-like receptor 2 also known as TLR2 is a protein that in humans is encoded by the TLR2 gene. TLR2 has also been designated as CD282 (cluster of differentiation 282). TLR2 is one of the toll-like receptors and plays a role in the immune system. TLR2 is a membrane protein, a receptor, which is expressed on the surface of certain cells and recognizes foreign substances and passes on appropriate signals to the cells of the immune system.26
TLR2 receptors are also expressed in leukocytes and endothelial cells.27
Our data demonstrate that TLR2 is the innate immune sensor for the S protein. Like other MyD88-dependent TLR pathways, ligation of TLR2 leads to the activation of transcription factors NF-κB and AP-1. Interestingly, while there was activation of NF-κB, AP-1 upstream signaling kinases such as ERK and JNK were not seen activated by S proteins. SARS-CoV-1 S protein activates the NF-κB pathway in human monocyte derived macrophages. COVID-19 patients also exhibited increased activation of the NF-κB pathway. Interestingly, in contrast to these findings, a separate study reported that SARS-CoV-1 S protein expressing baculovirus activates AP-1 but not NF-κB in A549 cells. Future studies dissecting the signaling pathway regulated by S protein of SARS-CoV-1 and CoV-2 may reveal further insight.
In summary, this study documents a potential mechanism for the inflammatory response induced by SARS-CoV-2. We demonstrate that SARS-CoV-2 S protein is a potent viral PAMP that upon sensing by TLR2 activates the NF-κB pathway, leading to the expression of inflammatory mediators in innate immune and epithelial cells. The effort so far in combating the COVID-19 pandemic is unprecedented, making it possible for the development of a number of vaccines within a year of outbreak. Since S protein is being targeted by most of the vaccine candidates, it is important to consider its inflammatory function in vaccine design. Considering the fact that new variants of SARS-CoV-2 with mutations in the S protein spread more easily and may confer more severe disease, the effectiveness of current vaccines remain uncertain. Thus, the importance of developing therapeutic drugs for COVID-19 remains high. This study suggests that TLR2 or its downstream signaling adapters could be therapeutically targeted to mitigate hyperinflammatory response in COVID-19 patients.
No conflict of interest declaration was provided.
Thank you for reading DoorlessCarp’s Scientific Literature Review. This post is public so feel free to share it.
KangAe Lee, Celeste M. Nelson, Chapter four - New Insights into the Regulation of Epithelial–Mesenchymal Transition and Tissue Fibrosis, Editor(s): Kwang W. Jeon, International Review of Cell and Molecular Biology, Academic Press, Volume 294, 2012, Pages 171-221, ISSN 1937-6448, ISBN 9780123943057,
Cameron AC, Touyz RM, Lang NN. Vascular Complications of Cancer Chemotherapy. Can J Cardiol. 2016 Jul;32(7):852-62. doi: 10.1016/j.cjca.2015.12.023. Epub 2015 Dec 28. PMID: 26968393; PMCID: PMC4989034.
Osiaevi I, Schulze A, Evers G, Harmening K, Vink H, Kümpers P, Mohr M, Rovas A. Persistent capillary rarefication in long COVID syndrome. Angiogenesis. 2022 Aug 11:1–9. doi: 10.1007/s10456-022-09850-9. Epub ahead of print. PMID: 35951203; PMCID: PMC9366128.
Østergaard L. SARS CoV-2 related microvascular damage and symptoms during and after COVID-19: Consequences of capillary transit-time changes, tissue hypoxia and inflammation. Physiol Rep. 2021 Feb;9(3):e14726. doi: 10.14814/phy2.14726. PMID: 33523608; PMCID: PMC7849453.
DoorlessCarp. Spike protein (inc vax) induced immunodeficiency & carcinogenesis megathread #29: The tryptophan catabolite or kynurenine pathway and long COVID. (2022)
Nissanka N, Moraes CT. Mitochondrial DNA damage and reactive oxygen species in neurodegenerative disease. FEBS Lett. 2018 Mar;592(5):728-742. doi: 10.1002/1873-3468.12956. Epub 2018 Jan 9. PMID: 29281123; PMCID: PMC6942696.
Bohlson SS, O'Conner SD, Hulsebus HJ, Ho MM, Fraser DA. Complement, c1q, and c1q-related molecules regulate macrophage polarization. Front Immunol. 2014 Aug 21;5:402. doi: 10.3389/fimmu.2014.00402. PMID: 25191325; PMCID: PMC4139736.
This is beyond helpful, it is revealing. Like most people I’ve been fascinated by long-Covid. Then I can across a study by Dr. Jackson Turner who found viral shedding 97 days after a positive test and long after remission. But how did the virus evade detection? Dr. Bruce Patterson provided the answer when he found it hiding inside monocytes and neutrophils which together with “profoundly diminished CD8 cells” explains a lot. The nail proteins on the virus act as a super-antigen that is carried around inside leukocytes which release fractilkine that disrupts the glycocalyx and allows the monocytes to stick to the blood vessels and disrupt NO. The Lymphopenia allows the neutrophil nets to build up and release elastin that causes the amyloidosis. And the monocytes carry that across the BBB causing brain fog and headaches. The gp120 proteins act as super-antigens in a chain reaction that results in mis-folded amyloids. Wow!
assuming the green banded pieces of text are taken from the study, i wonder why the researchers decided to include the pro vaccine propaganda about the emergency in german hospital due to rising numbers.
it probably has to do with its funding and getting it published as it seems that no single study is being performed and/or published that does not have some sort of knee jerk towards the big pharma backed "truth".
how about the researchers do the exact same study with people that never were infected with covid19 but were injected with the "vaccine"?
by way of retribution for having sold out to big pharma maybe?






















This is beyond helpful, it is revealing. Like most people I’ve been fascinated by long-Covid. Then I can across a study by Dr. Jackson Turner who found viral shedding 97 days after a positive test and long after remission. But how did the virus evade detection? Dr. Bruce Patterson provided the answer when he found it hiding inside monocytes and neutrophils which together with “profoundly diminished CD8 cells” explains a lot. The nail proteins on the virus act as a super-antigen that is carried around inside leukocytes which release fractilkine that disrupts the glycocalyx and allows the monocytes to stick to the blood vessels and disrupt NO. The Lymphopenia allows the neutrophil nets to build up and release elastin that causes the amyloidosis. And the monocytes carry that across the BBB causing brain fog and headaches. The gp120 proteins act as super-antigens in a chain reaction that results in mis-folded amyloids. Wow!
assuming the green banded pieces of text are taken from the study, i wonder why the researchers decided to include the pro vaccine propaganda about the emergency in german hospital due to rising numbers.
it probably has to do with its funding and getting it published as it seems that no single study is being performed and/or published that does not have some sort of knee jerk towards the big pharma backed "truth".
how about the researchers do the exact same study with people that never were infected with covid19 but were injected with the "vaccine"?
by way of retribution for having sold out to big pharma maybe?